
Viajemos a uno de mis periodos favoritos en la evolución humana: aquel en que algunos representantes de Homo tempranos comienzan a desarrollar comportamientos de cooperación. Se trata de grupos de pocas decenas de individuos y cierta división de tareas entre miembros. Es difícil pensar en el éxito de aquellos primeros humanos sin ciertas conductas complejas de organización. Aunque todas estas cuestiones siguen siendo objeto de debate, muchos me habréis visto defender en repetidas ocasiones que en este contexto tenemos que buscar también el origen de los cuidados entre humanos.
Manuel Domínguez-Rodrigo y colaboradores, grandes conocedores de la Garganta de Olduvai, presentan la evidencia más antigua conocida de aprovechamiento de un elefante, asociada al sitio EAK (Emiliano Aguirre Korongo), en el Lecho II de Olduvai, datada en unos 1,78 Ma (millones de años).
Se documenta un esqueleto parcial juvenil con 153 restos correspondientes a 46 elementos esqueletales de Elephas recki, junto con 80 artefactos líticos en el mismo nivel (herramientas de percusión, núcleos, lascas, subproductos de talla), bien conservados y con bordes afilados. Los autores interpretan esta acumulación como la concentración de un episodio de procesado de carcasa, no el resultado de una deposición casual. Huesos y artefactos forman una concentración muy densa en el centro del yacimiento, y el análisis espacial sugiere una asociación funcional entre ambos conjuntos.
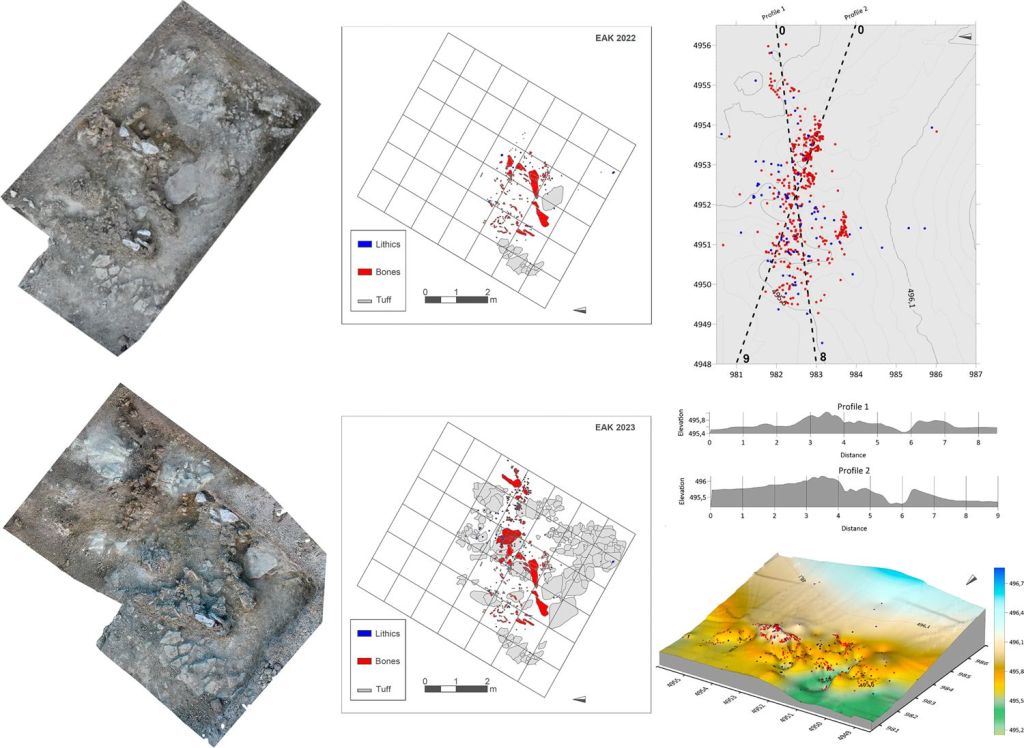
En general, la mera coexistencia espacial de herramientas y huesos no basta para demostrar una asociación causal, porque muchos yacimientos paleolíticos son palimpsestos formados por deposición sucesiva y por procesos postdeposicionales. Por ello, los autores combinan estudio tafonómico y espacial para buscar la asociación y descartar una coincidencia sedimentaria:
- Al menos dos huesos de elefante tienen marcas de rotura en fresco, que corresponderían a una acción intencional de homininos. No existe documentación de fracturas óseas similares realizadas por carnívoros.
- La distribución de los restos encaja con un procesado selectivo: los huesos y las herramientas aparecen concentrados donde cabría esperar el trabajo de descarnado y desmembramiento, no una dispersión aleatoria.
- Aunque la preservación es buena en muchas piezas, la cortical de varias estaba afectada por diagénesis, impidiendo detectar modificaciones superficiales.
- No se han encontrado marcas de corte sobre los restos óseos, aunque en el procesado masivo de grandes proboscídeos es raro encontrar marcas de corte, porque la desarticulación y el despiece pueden dejar pocas huellas visibles (Crader, 1983; Haynes y Krasinski, 2021).
Más allá del elefante de EAK, el estudio amplía el análisis a otros sectores de los lechos I y II de la garganta. A partir de 1,78 Ma aumentan las evidencias de explotación de megafauna, especialmente de proboscídeos e hipopótamos, en contextos que también muestran ocupaciones líticas más extensas y complejas:
- En la unidad inferior del Lecho II, la mayor concentración de restos de elefante e hipopótamo coincide con el área de máxima densidad de líticos del Lower Augitic Sands (LAS), la capa principal del Lecho II. Todos los conjuntos faunísticos de esa unidad salvo EAK se sitúan dentro de LAS. Aunque EAK está en la base del Lecho II, en la capa inmediatamente inferior a LAS, sigue su mismo patrón espacial.
- Mientras tanto, la unidad intermedia muestra una densidad más limitada espacialmente.
- La unidad superior vuelve a presentar un gran concentración alrededor los sitios de Bell Korongo (BK) y Specialist Cliff (SC), con abundantes restos de Sivatherium y Pelorovis asociados con altas densidades de herramientas.
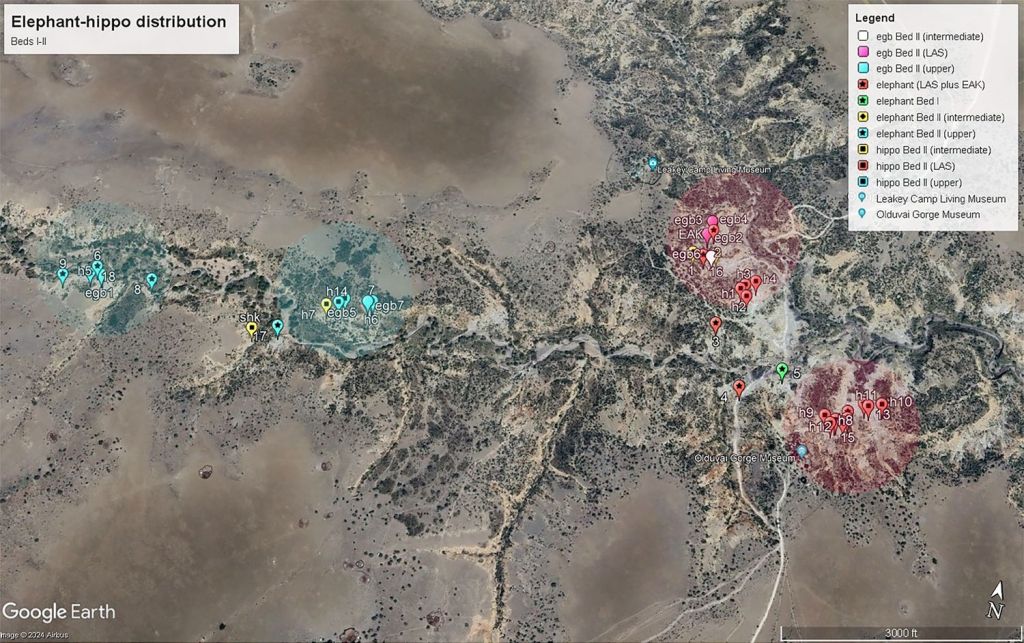
Esto lleva a los autores a proponer que la coincidencia espacial repetida entre megafauna y líticos a escala de paisaje no es aleatoria y parece reflejar la intervención de los homininos en varios momentos y sectores de Olduvai.
Aunque este hecho por sí solo no demuestra causalidad, sí constituye un marco espacial para situar el caso de EAK dentro de una pauta regional más amplia: la explotación regular de grandes carcasas no fue solo oportunista, sino una estrategia adaptativa vinculada a mayores necesidades energéticas, mayor tamaño de grupo y cambios tecnológicos cercanos al surgimiento del Achelense.
En este último punto, creo que hay que tomar con cautela una asociación explícita con el Achelense en esta reflexión, dado que el origen de este modo tecnológico se está adelantando con nuevas publicaciones, a la vez que va ampliándose la coexistencia con el Olduvayense y, con ello, retrasándose la desaparición de este último. La transición Olduvayense-Achelense es más compleja de lo que a veces estamos tentados a presentar.
En todo caso, tenemos nuevos datos que apoyan un umbral en torno a 2 Ma para el origen de los comportamientos humanos complejos relacionados con la cooperación social, la movilidad y la organización espacial-ecológica. Pero no olvidemos que muchas inferencias conductuales se basan en indicios como los patrones de fractura ósea y la superposición espacial en EAK, que deben seguir siendo completados con nuevos estudios.
Y si estabais esperando más carnaza sobre el debate caza vs. carroñeo, el artículo no propone que el acceso a la megafauna implicara una u otra conducta, pero sí menciona que desde muy temprano los homininos fueron capaces de intervenir de manera recurrente y estratégica sobre carcasas de gran tamaño. Podemos pensar, apoyándonos en publicaciones previas comparables y en modelos etnográficos y energéticos, que animales de este tamaño proporcionan alimento durante semanas a grupos relativamente numerosos (por ejemplo, de entre 20 y 50 homininos), lo que probablemente implicaría una cooperación social avanzada para dividir tareas, incluida la defensa de la carroña. Cada vez parece más obvio que los homininos empleaban distintas estrategias de obtención de recursos según las condiciones de cada escenario, lo que pudo contribuir de manera clave a su éxito adaptativo.
