El yacimiento de Prado Vargas, situado en el complejo kárstico de Ojo Guareña (Burgos, norte de la península ibérica), aporta información clave sobre las últimas poblaciones neandertales antes de la llegada de Homo sapiens a la región. Un estudio de carácter interdisciplinar (Navazo et al., 2021) reconstruye la vida cotidiana y las tradiciones culturales de estos grupos combinando análisis de estratigrafía, cronología, polen, fauna, industria lítica y restos humanos.
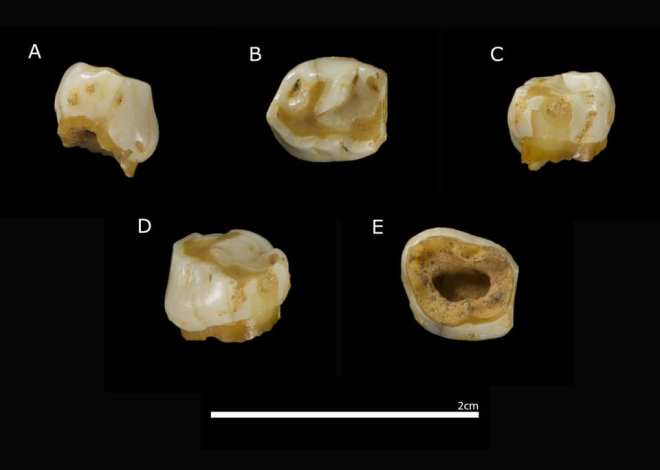
El nivel arqueológico más relevante, N4, se ha datado mediante OSL y radiocarbono entre 54,7 y 39,8 ka (miles de años), por tanto en el MIS 3, un periodo de intensos cambios climáticos. Este nivel se interpreta como un palimpsesto producto de ocupaciones recurrentes, probablemente estacionales (verano-otoño), cuando la cueva permanecía seca. Entre los hallazgos destaca un diente deciduo de un niño neandertal, evidencia directa de la presencia de individuos jóvenes en el grupo.
El diente de Prado Vargas: ¿de una niña o de un niño?
El mencionado diente es un deciduo inferior izquierdo (PV-1360) de un individuo de unos 9-10 años, que lo perdió de manera natural antes de la muerte. Inicialmente se le apodó Vera, que es el nombre de la nieta de Beni, dueño del prado donde se encuentra el yacimiento.
Su conservación ha permitido estudiar las proteínas del esmalte. El esmalte dental funciona como un sistema cerrado, protegiendo las proteínas originales durante decenas de milenios. Entre ellas destaca la amelogenina, codificada por dos genes parálogos: AMELX en el cromosoma X y AMELY en el cromosoma Y. Gracias a la espectrometría de masas aplicada a un pequeño fragmento de esmalte (unos 30 mg), fue posible detectar péptidos exclusivos de AMELY, lo que certifica que el individuo poseía un cariotipo XY. Es decir, cromosómicamente era un individuo masculino.
Este trabajo (Carrillo-Martín, 2025), presentado en el congreso de la ESEB (Evolutionary Society for Evolutionary Biology), constituye un nuevo ejemplo significativo de cómo la paleoproteómica puede suplir la ausencia de ADN en fósiles antiguos, aportando información biológica esencial, como la determinación del sexo o las relaciones filogenéticas.
Ahora bien, ¿cómo rebautizar al joven neandertal…? Yo propondría Vero, a partir del latín verus («verdadero»), para celebrar la determinación de su sexo cromosómico y, en breve, su certificación molecular como neandertal, gracias de nuevo a su proteoma.