Llevábamos algún tiempo sin volver al «mundo perdido» de Flores (Indonesia), una isla siempre separada del continente por un ancho brazo oceánico de decenas de kilómetros. A pesar de ello, dependiendo del periodo climático, esa distancia se reducía periódicamente a tal vez unos 20 km, lo que permitía que algunas especies alcanzaran la isla mediante eventos excepcionales de dispersión como, por ejemplo, balsas naturales empujadas por tormentas o tsunamis. Entre estas especies, llegarían grupos de homininos derivados de poblaciones tempranas de Homo, con algunos rasgos arcaicos que recuerdan incluso a Homo habilis. Las raíces ancestrales africanas de aquellas poblaciones posiblemente se acercaban a los dos millones de años (Ma). Sus descendientes ya habían dejado hace ~1,8 Ma una presencia antigua fuera de África de homininos con rasgos arcaicos, en Europa oriental (Dmanisi y Orozmani, Georgia) y tal vez en China central (Yunxian, con datación discutida).
En todo caso, algunos animales del sudeste asiático que llegaron a Flores, por un proceso llamado especiación alopátrica, impulsado por el aislamiento geográfico extremo, evolucionaron hacia especies endémicas como animales elefantes enanos (stegodones) o ratas gigantes de un metro (Papagomys armandvillei). Entre ellas, se incluyen los pequeños homininos encontrados en yacimientos como Liang Bua (donde se definió la especie Homo floresiensis) o Mata Menge. H. floresiensis era una especie de poco más de 1 m de estatura, unos 25 kg de peso y un cerebro de 400 cc, poco mayor que el de un chimpancé adulto actual. Su presencia en Liang Bua se ha datado en unos 190-50 ka. El hominino de Mata Menge, más antiguo que el de Liang Bua (~700 ka) también parece probablemente más pequeño. Por ahora, en espera de nuevos restos, suele asociarse al linaje floresiensis.
Un nuevo trabajo (Veatch et al., 2026) ha revisado de forma muy detallada los restos de stegodon de Liang Bua y concluye que no existe evidencia clara de una explotación sistemática de grandes presas ni de un uso habitual del fuego por parte de H. floresiensis, apoyándose en una combinación de tafonomía, zooarqueología cuantitativa, análisis experimental de marcas y comparación con restos quemados de roedores procedentes de niveles asociados tanto a dicha especie como a Homo sapiens.
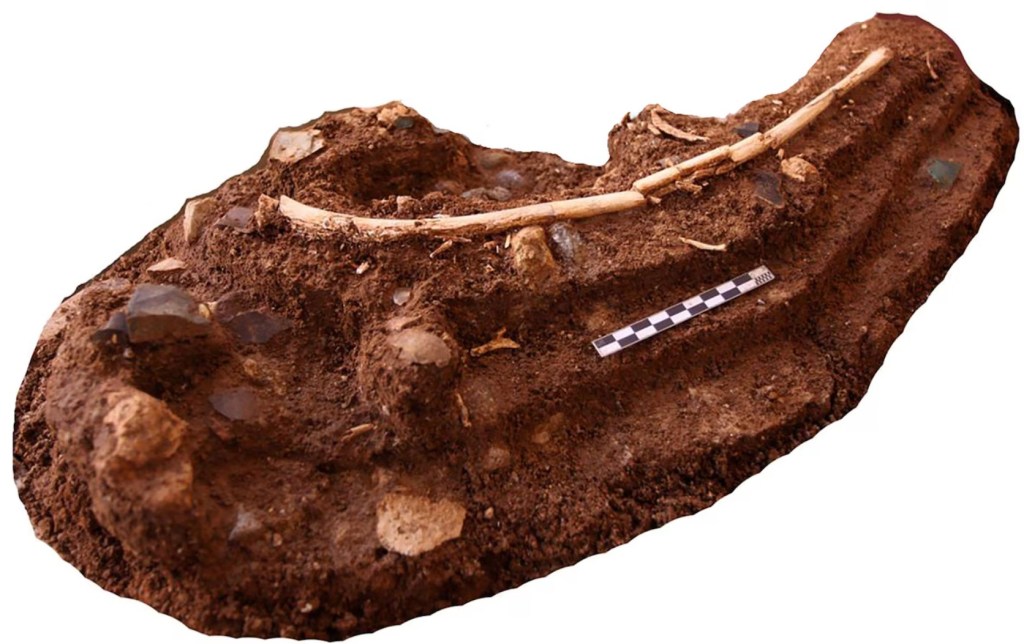
La muestra principal procede de las unidades 1 y 2 de Liang Bua, con presencia atribuida únicamente a H. floresiensis. Se estudiaron 3155 fragmentos de stegodon y 6906 elementos esqueléticos de múridos, muy fragmentados por causas posdeposicionales, y se reconstruyó tanto el contexto estratigráfico como la distribución espacial de los restos. Entre los restos de stegodon con edad identificable, 709 son de subadultos y 1511 de adultos, que corresponden a un mínimo de 9 individuos subadultos y 2 adultos.