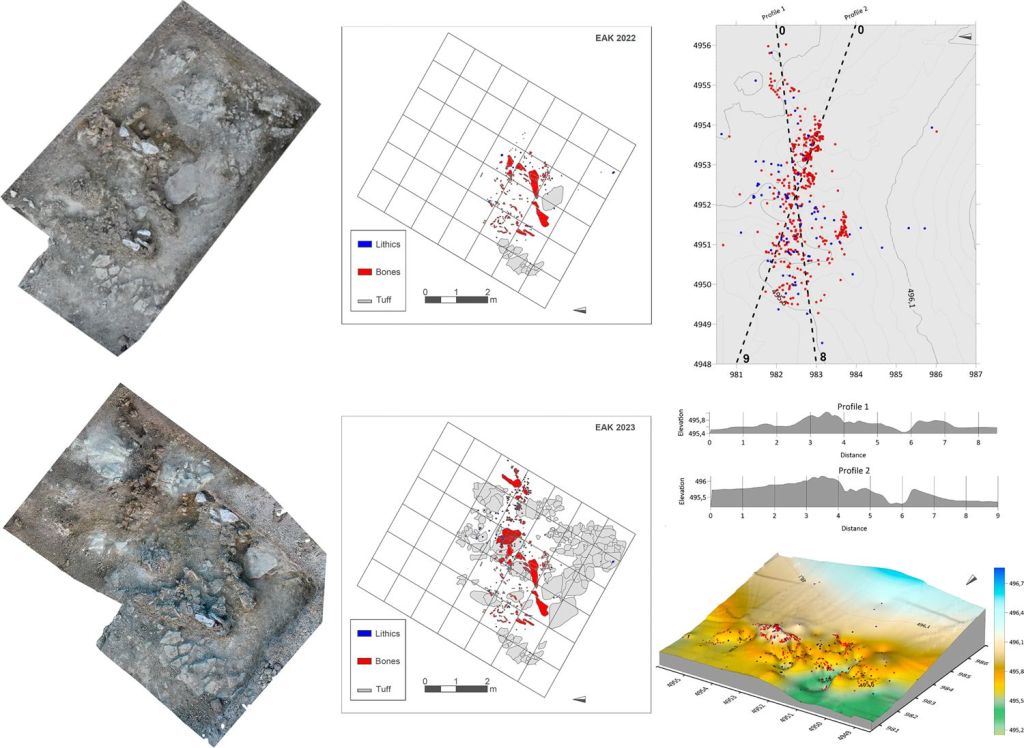
Un trabajo de Akbari, Perry, Barton y colaboradores (2026) observa un aumento en la intensidad de la selección natural en el registro genómico durante los últimos 10.000 años. Se trata del mayor estudio realizado hasta ahora sobre ADN humano antiguo, con datos de 15.836 euroasiáticos occidentales (10.016 con nuevos datos), procedentes de poblaciones antiguas de Europa y Oriente Próximo desde el surgimiento de la agricultura.
Los resultados muestran que la selección natural ha tenido un papel muy intenso en la evolución humana reciente, a través de numerosos casos de selección direccional, un proceso en el que variantes genéticas ventajosas aumentan rápidamente en frecuencia. Este trabajo aporta evidencia sólida de que la evolución humana no solo continúa, sino que se caracteriza por una mayor intensidad o detectabilidad de señales de selección en el registro reciente de Eurasia occidental.
El auge de la agricultura introdujo nuevos alimentos, patógenos y otros desafíos a medida que las poblaciones crecían y aumentaba la proximidad con animales domesticados. Esta gran transformación de los sistemas sociales y económicos humanos lógicamente tuvo consecuencias adaptativas. Sin embargo, los estudios genómicos hasta ahora solo habían identificado alrededor de una veintena de casos de selección reciente ampliamente aceptados, entre los que destacan:
- La persistencia de la lactasa, es decir, la tolerancia a la leche en adultos en buena parte de la población mundial, que es el caso paradigmático de selección fuerte y reciente.
- La pigmentación de piel, ojos y cabello como adaptación a la radiación ultravioleta.
- La resistencia a determinadas enfermedades infecciosas, como la malaria (anemia falciforme).
Otras posibles señales identificadas estaban más discutidas, como el metabolismo de la dieta (almidón, grasas, etc.) o ciertas funciones inmunitarias. En conjunto, predominaba una cierta idea de que los pulsos de selección fuerte y rápida eran relativamente escasos en la evolución humana reciente.
El nuevo estudio incrementa radicalmente el tamaño de la muestra y, sobre todo, permite analizar directamente cambios por selección direccional en la frecuencia de variantes genéticas en diferentes grupos que vivieron en diferentes épocas. Esto hace más posible distinguir la acción de la selección natural de otras fuerzas como la deriva genética, cambios aleatorios o reemplazos poblacionales que también alteran la composición genética (por ejemplo, la sustitución de cazadores-recolectores europeos por agricultores de Oriente Próximo), aunque asegurar esa distinción sigue siendo un trabajo complejo.
Como resultado, se identificaron 479 variantes candidatas con indicios de selección direccional, de distinta magnitud y robustez. Entre los principales patrones observados destacan:
- Enfermedades infecciosas y sistema inmunitario, gran protagonista de los resultados de este trabajo: variantes que afectan a la respuesta a patógenos y la inflamación, que sugieren una adaptación continua a epidemias y nuevos entornos, siempre teniendo en cuenta que las trayectorias de variantes inmunes suelen implicar combinaciones complejas, no adaptaciones lineales. Algunos ejemplos son:
- Una variante genética vinculada a un mayor riesgo de desarrollar esclerosis múltiple (Barrie et al. 2024) experimentó un aumento drástico en su frecuencia hace unos 6000 años, pero se ha vuelto menos común en algunos grupos europeos en los últimos 2000 años.
- Una variante vinculada a la susceptibilidad a la tuberculosis se volvió menos común en los últimos 3000 años (Kerner et al., 2021) pero, antes de esto, su frecuencia aumentó considerablemente, posiblemente debido a la aparición de otros patógenos.
- Una variante que confiere resistencia al VIH en los humanos modernos se hizo más común hace entre 6000 y 2000 años, posiblemente porque también protegía contra las bacterias causantes de la peste, aunque esta es una hipótesis muy debatida.
- Metabolismo y dieta: señales en genes implicados en el procesamiento de nutrientes y el uso de energía, más allá del caso clásico de la lactasa, que ilustran una adaptación compleja a dietas agrícolas.
- Rasgos complejos (poligénicos): combinaciones de alelos relacionados con rendimiento cognitivo, estatura o menor grasa corporal parecen haber aumentado su frecuencia, mientras que habrían disminuido la frecuencia de combinaciones relacionadas con diabetes tipo 2, esquizofrenia o trastorno bipolar. Estos resultados son inciertos, ya que los efectos poligénicos están calibrados en poblaciones modernas y son sensibles a sesgos poblacionales y estratificación, por lo que no está claro cómo se podrían extrapolar a fenotipos adaptativos en el pasado.
- Pigmentación: identificación de diez variantes asociadas a tonos de piel más claros y el cabello rojo.
- Otros rasgos fisiológicos: cambios en variantes relacionadas con crecimiento, desarrollo y funciones biológicas generales. Como curiosidad, una causa de la calvicie masculina se ha vuelto mucho menos común en los últimos 7000 años, lo que ha contribuido a una disminución estimada del 1-2 % en la prevalencia de la calvicie.
En conjunto, los resultados indican que la selección natural reciente ha sido mucho más frecuente y extendida de lo que se pensaba, afectando especialmente a rasgos complejos y poligénicos, no solo a mutaciones simples. Además, la intensidad de estos procesos parece tener un «pico» durante la Edad del Bronce, hace unos 5000 años, posiblemente como consecuencia de una intensificación de los cambios en el estilo de vida iniciados en el Neolítico, hace unos 10.000 años.




 hace 1,78 Ma: implicaciones para el origen de la cooperación")