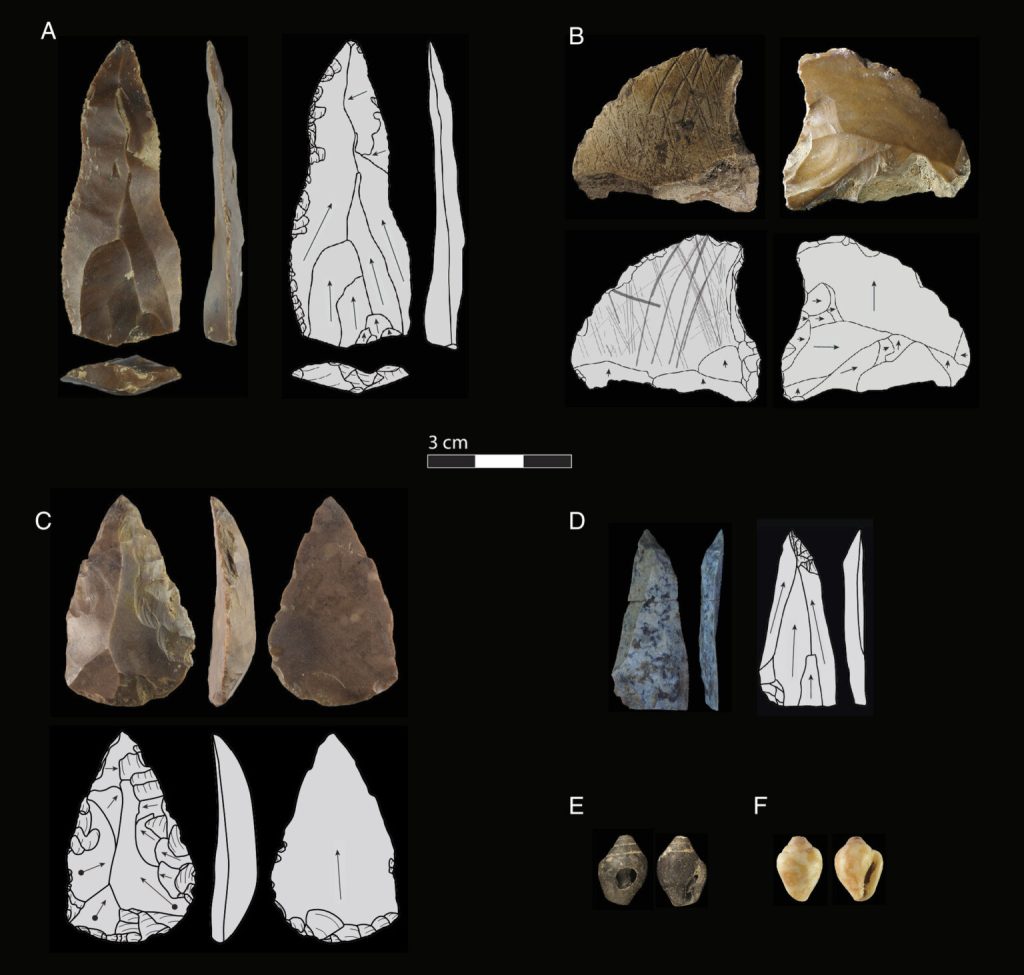
La cueva Üçağızlı II es un refugio natural en el sur de la actual Turquía. Un trabajo de Baykara y colaboradores documenta la ocupación secuencial de esta cueva por distintos grupos humanos durante unos 30.000 años: por neandertales desde 77 ka hasta 59 ka y, después, por humanos modernos desde 59 ka hasta 47 ka, sin que el registro arqueológico muestre una ruptura apreciable entre ambas ocupaciones.
Analizando más de 19.000 artefactos líticos recuperados en la cueva, los autores han encontrado que ambas especies emplearon las mismas técnicas de talla características del Paleolítico medio. Esta observación, junto con los restos de fauna y flora analizados, indica estrategias de subsistencia muy similares.
Además, se han encontrado 59 conchas de moluscos marinos (Columbella rustica) repartidas por varios niveles. Dado que este pequeño molusco carecía de relevancia por su valor alimenticio, se sugiere que las conchas se habrían seleccionado por su forma para usarlas en adornos personales. Este comportamiento no utilitario, probablemente relacionado con prácticas simbólicas, también muestra una continuidad cultural entre ambas especies.
Artefactos de la cueva de Üçağızlı II. Crédito: Baykara et al. (2026).
Como cada trimestre, aquí llega el resumen de las principales novedades publicadas en el ámbito de la paleoantropología, ordenadas cronológicamente según la antigüedad de los hallazgos estudiados.
Una nueva estimación para la divergencia de los linajes chimpancé y humano la sitúa en 8,69-7,28 Ma, anterior a los c. 7 Ma que se venía asumiendo, basada en datos genéticos y evidencias fósiles (d’Oliveira Coelho et al., 2026).
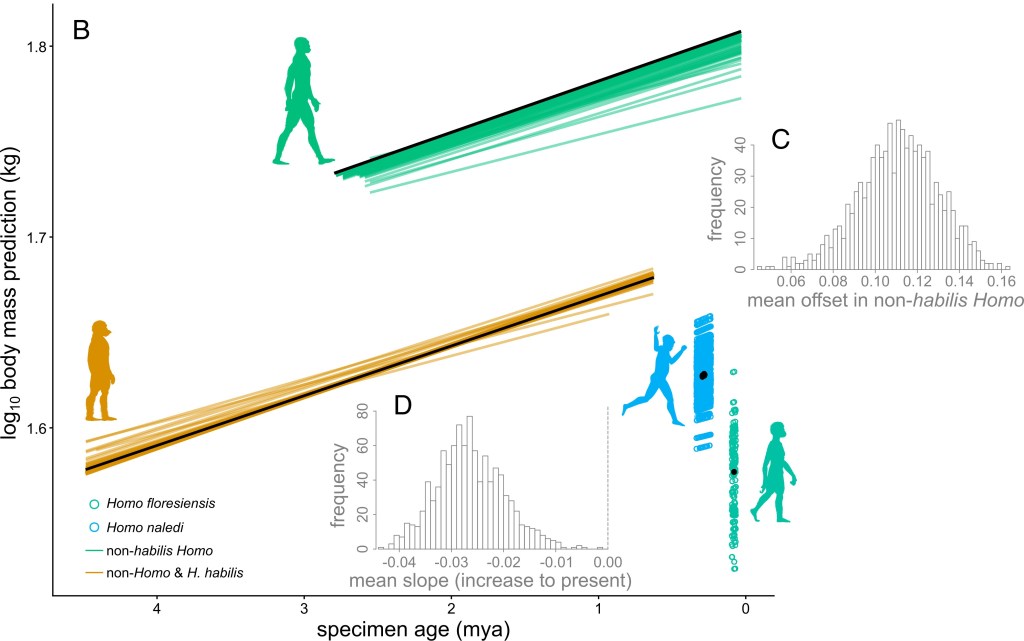
A partir de un análisis de 386 especímenes de 21 taxones, Gardner et al. (2026) publican evidencias de un fuerte aumento de la masa corporal para todas las especies tardías de Homo, excepto Homo habilis, no como tendencia continua, sino como un aumento progresivo en los homininos más antiguos, seguido de un cambio concentrado dentro del género Homo, acompañado de ramas paralelas que siguieron caminos distintos.
Evolución del tamaño corporal en Gardner et al. (2026)
Hunter y colaboradores han estudiado la variación en la morfología carpiana de los homininos, un rasgo relevante desde el punto de vista biomecánico, encontrando una retención de rasgos potencialmente ancestrales que persistieron hasta Homo naledi. Con ello, sugieren que el carpo de los homininos habría evolucionado a partir de una muñeca similar a la de los simios africanos, y que su reorganización relacionada con la manipulación de herramientas de piedra habría ocurrido ya avanzado el Pleistoceno medio. Además, proponen que el último ancestro común chimpancé-humanos caminaría con nudilleo (knuckle-walking).
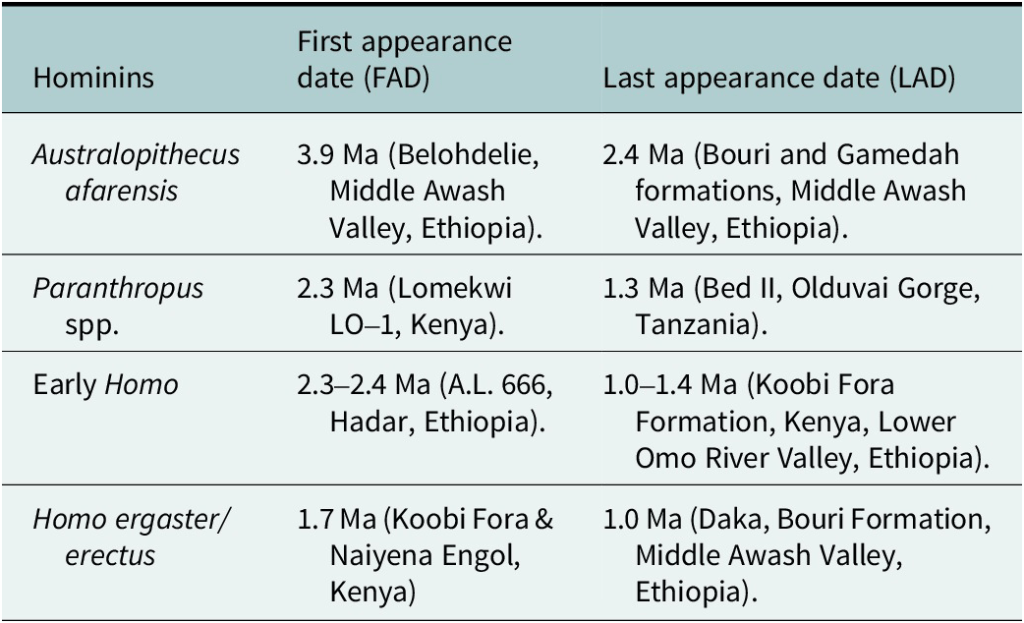
A partir de evidencias de útiles líticos asociadas con la desaparición de especies de Australopithecus, Paranthropus y Homo temprano en el registro africano oriental entre 0,8 y 3,5 Ma, Shea (2026) sugiere que la transición evolutiva entre los primeros Homo y Homo ergaster/erectus parece estar correlacionada con un cambio en la talla de ocasional a habitual, pero las últimas apariciones de Australopithecus y Paranthropus no parecen estar correlacionadas con cambios importantes en el registro.
Primera y última aparición de homininos del Plio-Pleistoceno en África oriental (Tabla 2 en Shea, 2026).
Los nuevos hallazgos de restos fósiles de las extremidades inferiores de parántropos y australopitecos han permitido comparar su estructura ósea interna, concretamente entre especímenes de Paranthropus robustus de Swartkrans (parte de la pelvis, un fémur casi completo y una tibia completa del individuo SWT1/HR-2 del Miembro 1 de Swartkrans) y de especímenes de Australopithecus del Miembro 4 de Sterkfontein (Cazenave et al., 2026). La estructura ósea en los parántropos se acerca más a la de los grandes simios africanos que a la de los australopitecos. Esto sugiere una flexión más frecuente en P. robustus de las articulaciones del tobillo, la rodilla y la cadera, normalmente asociada a la trepa. Los autores interpretan una bipedación terrestre más constante en Australopithecus, y un repertorio de locomoción más diverso en los posteriores P. robustus. Como conclusiones derivadas, ambos taxones explotarían nichos ecológicos diferentes (los parántropos, un territorio más extenso). Nuevamente, se observa una evolución nada lineal de la bipedación en los homininos, y surge la necesidad de comparar este comportamiento locomotor con el de Paranthropus boisei en África oriental, para buscar similitudes que podrían haber caracterizado al género en cuanto a la frecuencia de posturas muy flexionadas de la cadera, la rodilla y el tobillo.
Hasta ahora, era habitual pensar que los neandertales tardíos vivían en poblaciones pequeñas, aisladas y con claros signos de endogamia. Suelo contar esto diciendo que los Homo sapiens con quienes se iban cruzando tan solo les dieron el golpe definitivo a unas poblaciones ya debilitadas demográficamente. Esta idea se apoyaba principalmente en los genomas de varios neandertales de cuevas de los montes Altái (Siberia), como Denisova 5 (120 ka) y Chagyrskaya 8 (~80 ka), complementados por otros individuos como Okladnikov 2 (45-50 ka), Chagyrskaya 84 (~80 ka) y otros con menor cobertura. A ello se suma la hipótesis de la asimilación, según la cual algunos individuos o pequeños grupos terminaron integrándose (también genéticamente) en poblaciones sapiens.
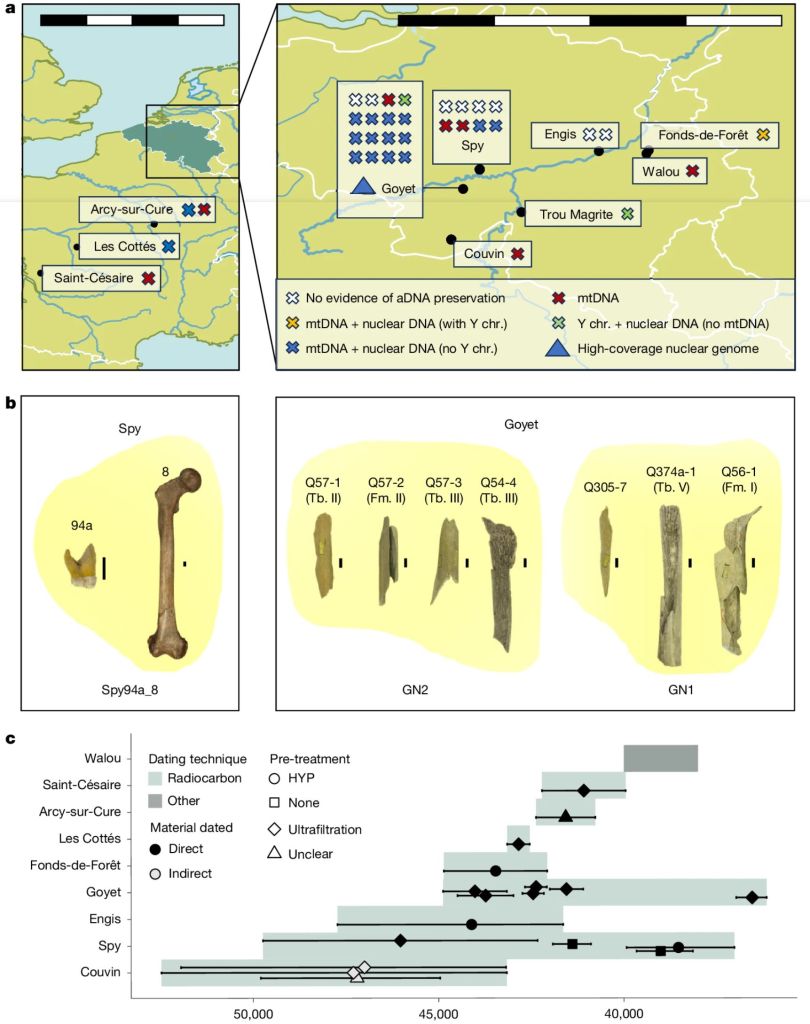
Un nuevo trabajo de Bossoms Mesa y colaboradores matiza la propuesta del deterioro genético. El estudio abarca el ADN de 27 neandertales procedentes de diez yacimientos de Bélgica y Francia de los últimos 52,5 ka, incluyendo GN1 (Goyet), Spy94a, Z4-1514 (Les Cottés), Fond-de-Forêt 1, Trou Magrite 2423-36, entre otros. Por una parte, se han usado nuevas técnicas de descontaminación, extracción, preparación de bibliotecas genéticas y captura selectiva de fragmentos de ADN neandertal. Por otra, se ha secuenciado el quinto genoma neandertal de alta cobertura (es decir, leyendo repetidamente cada posición del ADN para minimizar errores) publicado hasta la fecha, tras los genomas de Denisova 5 (Siberia, Rusia), Vindija 33.19 (Croacia), Chagyrskaya 8 (Siberia, Rusia) y Mezmaiskaya 1 (Cáucaso, Rusia).
Uno de los indicadores genéticos más informativos se ha basado en los llamados tramos de homocigosidad o ROH (Runs of Homozygosity). Cada persona hereda una copia de cada cromosoma del padre y otra de la madre. Cuando ambos progenitores son familiares cercanos, es mucho más probable que transmitan exactamente las mismas variantes genéticas heredadas de un antepasado común. El resultado son largas regiones continuas del genoma en las que ambas copias son idénticas. Cuanto más largos y numerosos son esos tramos, más reciente e intensa ha sido la consanguinidad. Esos largos ROH en los neandertales asiáticos son los que principalmente habían conducido a la interpretación de la endogamia. Sin embargo, los neandertales tardíos del noroeste europeo no presentan apenas esos largos tramos, lo que implica que no constituían grupos pequeños aislados ni una población uniforme genéticamente, sino una metapoblación regional formada por comunidades conectadas mediante flujo génico, dentro de la cual persistieron distintos linajes durante decenas de miles de años. Estas comunidades estaban más emparentadas entre sí que con otros neandertales contemporáneos de Europa.
Yacimientos, restos esqueléticos y cronologías en el estudio de Bossoms Mesa et al. (2026)
Recordemos la enigmática especie Homo naledi, de tamaño sorprendentemente pequeño, conocida solo en el sistema de cuevas sudafricanas de Rising Star. Este grupo humano existió hace unos 300.000 años, de forma casi contemporánea a la evolución de Homo sapiens en regiones no muy lejanas. Madupe et al. (2026) han estudiado la proteína amelogenina de 23 dientes de H. naledi, pertenecientes a un mínimo de 20 individuos. Esta proteína tiene variantes específicas diferentes según provenga de un gen del cromosoma X femenino o de un gen del cromosoma Y masculino. Los autores han hallado que, en 19 de los 20 individuos la variante AMELY (macho) estaba ausente (probabilidad >95%), y también lo estaba probablemente (50-95%) en el otro. Por tanto, entre 10 individuos adultos y 10 subadultos analizados, aparentemente todos son hembras.
Este es un hallazgo totalmente inesperado. Para que nos hagamos una idea, en una muestra de 20 individuos distintos, asumiendo una proporción sexual poblacional de 1:1, la probabilidad de encontrar todos los individuos hembra es extremadamente baja, 0,0000954%.
Por tanto, el estudio a mí me genera muchísimas preguntas que deberán ser trabajadas a partir de ahora:
Tenemos que revisar cualquier interpretación previa respecto al leve dimorfismo sexual que se había predicho entre machos y hembras de la especie, atendiendo a la distinción entre el tamaño de los esqueletos. Destaca el espécimen U.W. 102a (individuo LES1, apodado Neo), considerado tradicionalmente uno de los mejores candidatos a macho por su robustez, tamaño corporal y craneal. Pues bien, este individuo también carece de AMELY, es hembra.
Nos sorprendía el pequeño tamaño de estos humanos (del orden de 150 cm y 45 kg), resultando ahora que todos los especímenes de la muestra analizada son hembras. Hemos de esperar a la aparición de machos de H. naledi para revisitar nuestras interpretaciones del tamaño corporal. Podemos incluso especular con un tamaño corporal de machos y un dimorfismo sexual tal vez más próximo al de los humanos modernos que estaban evolucionando no lejos del sistema Rising Star.
¿Puede corresponder este hallazgo a una motivación económica, el reparto de tareas entre machos y hembras? Para ello, debemos asumir que las hembras eran más pequeñas y podían acceder con menos dificultad por las galerías de Rising Star para realizar alguna actividad determinada. Sin embargo, el tamaño de los subadultos de ambos sexos no excedería el de una hembra adulta, lo que debilita esta hipótesis.
¿Puede corresponder a una motivación cultural, una práctica funeraria por la cual depositaron solo cuerpos de mujeres en las cuevas? Ni siquiera hay bebés de sexo masculino que habrían sido enterrados con sus madres. Este escenario cultural reforzaría la hipótesis funeraria en H. naledi que ha sido cuestionada o, más bien, ilustraría una práctica funeraria muy concreta.
En el estudio se menciona como referencia la proporción de sexos desequilibrada 1:0,5 (F:M) en los individuos de la cueva portuguesa del Escoural (Granja et al., 2023), pero este caso es del Neolítico.
A mí me gusta más pensar en la Sima de los Huesos (España) que, con 430 ka de antigüedad, es un caso mucho más próximo cronológicamente al de H. naledi. Entre los 29 individuos de la Sima, el sesgo existente es el fuerte predominio de adolescentes y adultos jóvenes, que representan a más del 80%. En cambio, no hay sesgo por sexos en la Sima. Este caso suele considerarse un depósito intencional, con el que a veces se ha comparado a Rising Star.
Sin tener machos, tampoco podemos apresurarnos a buscar encajes de la escasa variabilidad morfológica de H. naledi comparándola con la de otras especies humanas. Además, H. naledi presenta dos sustituciones de aminoácidos: una variante derivada en la amelogenina X (AMELX) respecto a los demás representantes conocidos del género Homo y una variante arcaica P635 en COL17A1 (una proteína estructural presente en las células que forman el esmalte), identificada en cinco individuos. Esta última también aparece en dos ejemplares de Paranthropus robustus (SK835 y SK14132), mientras que humanos modernos, neandertales y denisovanos presentan la variante derivada A635. Más incógnitas a despejar.
Además, no hay variación en las secuencias de proteínas analizadas. Todos los individuos tenían secuencias muy similares. Esto podría indicar el aislamiento de esta población humana y/o su descendencia de una población fundadora muy pequeña. Una población grande y diversa es inconsistente con estos resultados.
Las conclusiones de este trabajo se verían alteradas si se demostrara que algunos machos de H. naledi presentaban una deleción o una mutación del gen AMELY que impidiera su detección mediante paleoproteómica. Los autores consideran poco probable este escenario y descartan que la ausencia de AMELY se deba simplemente a limitaciones técnicas o a una mala conservación de la proteína, ya que en otros fósiles de Australopithecus africanus, Paranthropus robustus y Homo antecessor sí se han identificado con éxito individuos masculinos mediante este mismo procedimiento.
El uso del fuego ya estaba documentado en la cueva sudafricana de Wonderwerk hace 1 millón de años (Ma), cuando un estudio (Berna et al., 2012) identificó restos de vegetales y huesos con huellas térmicas en el estrato 10. Dado que este hallazgo tuvo lugar a unos 30 metros de la entrada de la cueva, es difícil que su ubicación pueda explicarse por restos de incendios naturales llegados accidentalmente, sino que es más probable la intervención humana. Por otra parte, esta cueva es conocida también por la excelente conservación de los sedimentos y los restos arqueológicos, en una secuencia estratigráfica prácticamente continua que abarca desde el primer Olduvayense (1,93 Ma) hasta el Achelense plenamente desarrollado (1,07 Ma). Además, allí se ha documentado la ocupación humana más antigua conocida en el interior de una cueva (Shaar et al., 2021).
Como de costumbre, en la evolución humana debemos tener en mente las distintas etapas en el proceso gradual de nuestra relación con el fuego: la observación de incendios naturales y el aprovechamiento oportunista que comenzaron a hacer algunos homininos hace casi 2 Ma, el transporte de brasas, el mantenimiento deliberado de hogueras, la producción controlada del fuego, y el desarrollo de técnicas más diversas y específicas para un uso sistemático. Sin embargo, determinar los umbrales de cada etapa resulta extraordinariamente difícil. La conservación de las evidencias es escasa y es un reto distinguir entre restos de incendios naturales aprovechados, fuegos mantenidos o transportados, y creación de fuegos por homininos. Asimismo, hay que tener en cuenta que estas capacidades se desarrollarían en momentos distintos en cada región geográfica.
Infografía con las etapas de uso, mantenimiento y control del fuego en la evolución humana. Crédito: Roberto Sáez
Un nuevo estudio (Marín-Monfort et al., 2026) revela signos de uso recurrente del fuego en los estratos 11 y 12 de Wonderwerk, fechados entre 1,07 y 1,79 Ma, según se ha observado en miles de fragmentos óseos de micromamíferos con alteraciones térmicas. Se han reconocido al menos dos concentraciones diferenciadas de estos restos, separadas varios metros entre sí, que los autores interpretan como posibles focos de combustión. Además de por su antigüedad, el hallazgo destaca por su recurrencia: los indicios aparecen en niveles estratigráficos separados por largos intervalos temporales, sugiriendo que el transporte y mantenimiento del fuego no eran ocasionales, sino que formaban parte del repertorio conductual habitual en aquel lugar. Las primeras cronologías de estos depósitos, en los niveles más antiguos de la secuencia, coincidirían con una fase muy temprana de Homo erectus en la región, donde esta especie aparece al menos hace 2 Ma (el cráneo parcial DNH 134 de Drimolen).