
Actualizado marzo 2026
La mandíbula SK 15 fue descubierta en 1949 en Swartkrans Member 2 y está datada en unos 1,4 Ma (millones de años). Aunque presenta cierta distorsión en su lado derecho, es una de las mandíbulas homininas mejor conservadas y completas del registro paleoantropológico africano. Su buena conservación permite observar un cuerpo mandibular bajo y grueso, comparable al de algunos especímenes de Homo de antigüedad similar, aunque más pequeño en todas sus dimensiones que el de Paranthropus robustus. Los molares de SK 15 muestran un patrón de aumento distal en el diámetro mesiodistal (M1<M2<M3), característica común en los australopitecos y algunos Homo tempranos, pero rara en H. erectus.
Este fósil se atribuyó originalmente a Telanthropus capensis, una denominación introducida para describir formas intermedias entre los autralopitecos y Homo erectus/ergaster, pero actualmente en desuso. Desde la década de 1960, se ha asignado a H. ergaster, junto con SK 45 y SK 847, dos fósiles de Swartkrans Member 1, cuya datación es más antigua, de 2,22 Ma. No obstante, las asignaciones taxonómicas son complicadas. SK 847 es un cráneo fragmentario con maxilar casi edéntulo, mientras que SK 45 es un fragmento de hemimandíbula con dos molares muy desgastados. Además, tampoco es posible comparar SK 15 con el cráneo DNH 134 de Drimolen, que es el Homo erectus más antiguo conocido, ya que carece de dentición. Este registro fósil limitado y confuso de Homo ha llevado a que SK 15 se haya relacionado sorprendentemente con tres géneros de homininos: Australopithecus, Paranthropus y Homo (Homo habilis, Homo gautengensis y Homo naledi). Incluso las características dentales llevaron a los primeros investigadores a compararla con la mandíbula de Mauer (H. heidelbergensis).
En un nuevo trabajo, Zanolli y colaboradores han reestudiado SK 15 mediante microtomografía de rayos X, modelado 3D y análisis morfométricos geométricos. Según este estudio, la forma de la sínfisis mandibular es similar a la de los australopitecos, mientras que la arcada dental y la unión esmalte-dentina se aproximan a las de los parántropos. Básandose en estas observaciones, los autores concluyen que SK 15 perteneció a Paranthropus. Sin embargo, presenta características que lo diferencian de las especies conocidas de este género (P. aethiopicus, P. boisei y P. robustus), como su menor tamaño y la gracilidad del hueso, en comparación con las mandíbulas de P. robustus de Swartkrans, así como la ausencia en SK 15 de algunos rasgos dentales muy características de otros parántropos, tales como un protostílido desarrollado (una cúspide bien formada), un M3 que se estrecha distalmente y raíces molares cortas.
Estas diferencias han llevado a los autores a proponer una nueva especie, Paranthropus capensis, la primera definida en este género desde P. aethiopicus (Arambourg y Coppens, 1967). Paranthropus viene a significar «cerca del hombre», reflejando su posición lateral en la filogenia humana, mientras que capensis alude a su procedencia sudafricana. Clarke y Pickering (2026) refuerzan esta propuesta de eliminación de SK 15 del género Homo y atribución a P. capensis.
La divergencia de P. capensis respecto a P. robustus se habría producido hace más de 2 Ma, en un periodo de gran variabilidad morfológica dentro de los parántropos. Posteriormente, varias especies de este género habrían coexistido hace 1,4 Ma, ocupando nichos ecológicos distintos: mientras que P. robustus estaría adaptado a dietas basadas en materiales vegetales duros, la gracilidad de P. capensis indica hábitos alimentarios diferentes.
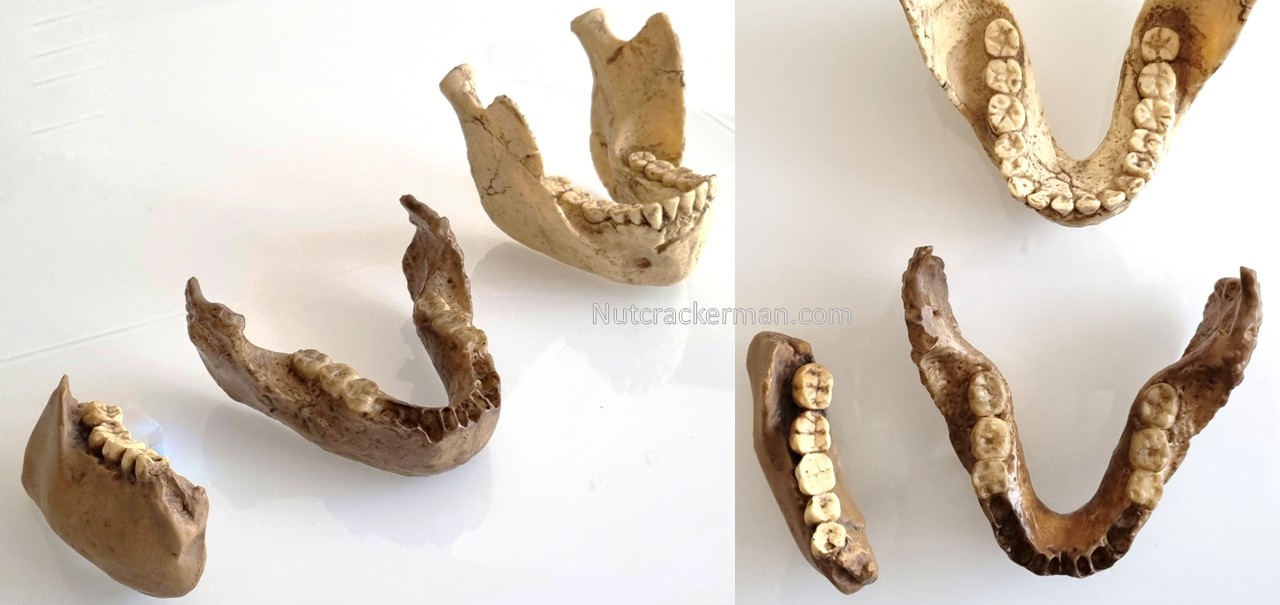
Comparación de: [centro] SK 15 Paranthropus capensis (reconstrucción 2); [izquierda] TM 1517 holotipo de P. robustus; [derecha/arriba] SK 23 P. robustus (reconstrucción). Los molares son comparables, pero el cuerpo mandibular es más bajo y grácil en P. capensis. Crédito: R. Sáez.
El registro fósil del sur de África entre el Plioceno final y el Pleistoceno temprano refleja una notable diversidad de homininos, incluyendo tres posibles especies de Australopithecus (Au. africanus, Au. sediba y Au. prometheus) y, posiblemente, dos o tres especies de Homo temprano (H. erectus, como el mencionado DNH 134; H. gautentensis, siendo su holotipo Stw 53 y 17 paratipos, propuesta por D. Curnoe en 2015, y tal vez H. habilis, con presencia nada clara en esta región; a ellas se añade H. naledi en el Pleistoceno medio). Dentro del genéro Paranthropus, se han identificado al menos dos especies en la región: P. robustus y la recién propuesta P. capensis. Los autores de esta última sugieren la reevaluación de otros especímenes de Swartkrans, Drimolen y Kromdraai, clasificados hasta ahora como P. robustus, alguno de los cuales podría ser P. capensis.
Además, existe un fósil sorprendente identificado como P. boisei (Skinner et al., 2018), siendo ésta una especie conocida únicamente en el este africano. Se trata de un segundo molar enorme, Gondolin GDA 2. Sin embargo, su tamaño también podría situarse dentro de la variabilidad de P. robustus, considerando que la distribución de tamaños en el registro de esta especie procedente de Swartkrans y Drimolen parece estar sesgada hacia una mayor proporción de ejemplares jóvenes y adultos más pequeños (Grine et al., 2012).

Comparación de SK 15 Paranthropus capensis [centro] con dos especímenes de Homo temprano esteafricanos: [izquierda] KNM-ER 992, holotipo de Homo ergaster, con cuerpo más grácil y dientes más pequeños y de estructura más simple; [derecha] KNM-ER 1802, Homo rudolfensis, con mandíbula robusta, molares con grandes coronas y raíces complejas. Crédito: R. Sáez
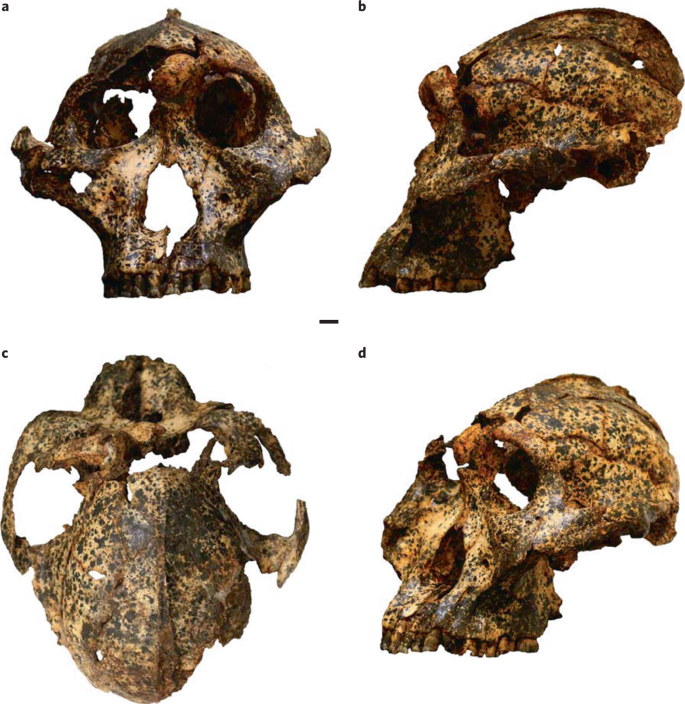
La diversidad taxonómica dentro de Paranthropus en esta región también se amplía con otra propuesta reciente que clasifica los fósiles de Drimolen dentro una subespecie de P. robustus: P. robustus ukusa, (J. M. Martin et al., 2024). Su holotipo es el cráneo DNH 155, acompañado por los paratipos los cráneos DNH 7 y DNH 152. P. robustus ukusa sería anterior al resto de parántropos de la región (P. robustus robustus) y se diferenciaría de ellos por una raíz cigomática más posterior respecto a la hilera dental, un hueso petroso orientado más sagitalmente, un extremo supraorbital redondeado en vez de cuadrado, una cresta sagital que comienza en los parietales en lugar del frontal, y menor separación cigomaticomaxilar. Este cambio evolutivo en el linaje habría sido impulsado por la selección sobre el sistema masticatorio, en respuesta a presiones dietéticas asociadas a un entorno cambiante.
Como siguientes pasos, sería recomendable un reanálisis exhaustivo de los especímenes provenientes de las cuevas sudafricanas para comprobar si se consolida P. capensis como una especie diferenciada. La diversidad de grupos de parántropos que se está proponiendo para el sur de África parece excesivamente ambiciosa para un registro fósil tan limitado, incluida la presencia de dos subespecies contemporáneas de P. robustus. Por otro lado, la escasa representación de Homo tempranos en este registro resulta intrigante y podría deberse a una clasificación errónea de ciertos fósiles. Tal vez cabe plantearse la reflexión contraria para el este africano, donde hay una multiplicidad de formas de Homo temprano, incluidos pequeños habilinos parecidos a australopitecos, o los robustos y enigmáticos H. rudolfensis.

Gracias Roberto.
Parece que una vez más el linaje no parece claro, y el escaso número de restos no aporta luz.
Saludos.
Me gustaLe gusta a 1 persona