
No es novedad que el paradigma del siglo XXI en evolución humana sea un rompecabezas genético reconstruido a partir del ADN antiguo (ADNa). En el último mes, dos estudios amplían nuestro conocimiento y proponen hipótesis tentativas (siempre con la cautela de que son modelos para avanzar, no conclusiones definitivas).
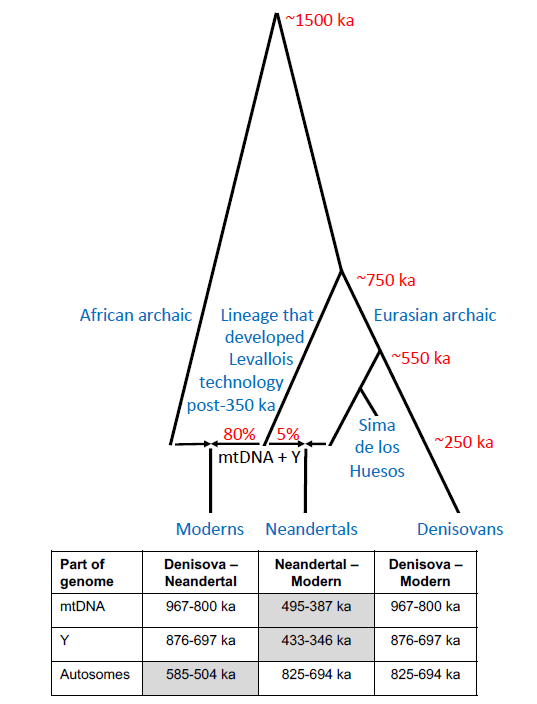
Un preprint del genetista David Reich (2026) propone un modelo demográfico para explicar aspectos de la historia de los linajes de neandertales y de humanos modernos. Según este modelo, hace entre 400 ka y 250 ka (miles de años) se produjo una expansión demográfica africana (expansión de rango o range expansion) de poblaciones asociadas a la Middle Stone Age (MSA) y la tecnología de núcleo preparado Levallois. Al expandirse hacia Eurasia, estas poblaciones habrían interactuado con grupos locales, probablemente similares a los representados en la Sima de los Huesos (Atapuerca, España), produciendo una mezcla genética muy asimétrica: la población resultante tendría aproximadamente 95% de ascendencia local basal (tipo Sima) y solo 5% de ascendencia moderna en ADN nuclear.
Aunque sería limitada esa contribución autosómica (a través de los autosomas, cromosomas no sexuales), el modelo de Reich muestra que bastaría para explicar el reemplazo completo de linajes uniparentales, como el ADN mitocondrial (ADNmt) y el cromosoma Y, debido a la dinámica de deriva y a redes de parentesco estructuradas. Las simulaciones indican que incluso una expansión numerosa puede dejar únicamente ese 5% de ascendencia autosómica africana en la población resultante. Este escenario corresponde cronológicamente a la aparición de paralelos culturales entre África y Eurasia en el Paleolítico medio, como la tecnología Levallois, el hafting (enmangado de herramientas) más sofisticado o el uso intensivo del fuego.
Este estudio puede vincularse con otra publicación reciente sobre el flujo génico africano hacia neandertales alrededor de 250 ka, con posibles sesgos sexuales en la mezcla genética. Platt y colaboradores (2026) han detectado un exceso de ascendencia africana en el cromosoma X de los neandertales, patrón que refleja a su vez el déficit de ADN neandertal en el cromosoma X de los humanos modernos y la ausencia de ADNmt neandertal en estos, tras las hibridaciones ocurridas hace unos 50 ka. Este patrón se ha interpretado como indicio de posibles sesgos sexuales en la mezcla genética, compatibles con la integración de mujeres sapiens africanas en poblaciones neandertales locales. No obstante, este resultado no excluye que parte del déficit de ADN neandertal en el cromosoma X observado en humanos modernos también esté relacionado con procesos de selección negativa contra variantes perjudiciales.
Así, la hipótesis de dispersión matrilineal basada en el exceso de cromosoma X sugiere que redes sociales de parentesco maternas habrían facilitado la expansión geográfica de grupos humanos mientras se incorporaban alelos potencialmente adaptativos de poblaciones locales, es decir, variantes genéticas que mejorarían la adaptación a ambientes concretos. En ese contexto, los linajes de ADNmt y cromosoma Y de origen africano podrían haberse extendido ampliamente incluso con una mezcla genética limitada.
Llama la atención la posible relación directa que se plantea entre la expansión africana y la difusión de la tecnología Levallois. Los denisovanos también utilizaron tecnologías de núcleo preparado similares, y recordemos que la divergencia genética entre denisovanos y neandertales se sitúa en torno a 585-504 ka. Estas tecnologías se han identificado en la cueva Denisova (250-200 ka), en yacimientos del este asiático (Guanyindong o Panxian Dadong, ~180 ka) y sur asiático (Attirampakkam, ~385 ka). Por tanto, la difusión de la tecnología de núcleo preparado pudo haberse producido mediante procesos culturales o evoluciones paralelas, sin necesidad de vincularla a la expansión biológica africana (Hawks, 2026).
Esta discusión también recuerda la llamada hipótesis del Modo 3 propuesta por Foley y Lahr (1997), según la cual las tecnologías de núcleo preparado del Paleolítico medio (Modo 3/Levallois/MSA) se originaron en África y se expandieron por el oeste de Eurasia, junto con una dispersión de poblaciones en momentos cálidos que habría contribuido a la posterior diferenciación regional de los linajes sapiens y neandertales. Estas tecnologías no deben interpretarse como indicadores directos de «modernidad cognitiva», sino como parte de un complejo proceso demográfico y cultural en el Pleistoceno medio.
Otros factores demográficos también pudieron influir en los patrones genéticos observados. Por ejemplo, la discordancia entre el genoma nuclear y los linajes uniparentales podría explicarse mediante deriva genética o hitchhiking (mayor frecuencia en un alelo neutro o ligeramente perjudicial por estar ligado a un gen beneficioso que tiene selección positiva) en poblaciones conectadas, como serían los primeros grupos sapiens y neandertales que tuvieron encuentros esporádicos (Hawks, 2026). Por otra parte, estudios sobre la estructura genética africana profunda muestran que las poblaciones africanas ya presentaban una subestructura compleja hace ~300 ka (Cousins et al., 2025), con ~80% de linajes relacionados con el clado neandertal-denisovano posteriores a 750 ka y ~20% de linajes muy antiguos o «fantasma» que divergieron hace ~1,5 Ma (millones de años), reflejando mezcla continua y compleja entre múltiples linajes arcaicos.
Por tanto, esta complejidad genética africana anterior a 300 ka indica que la población MSA no era homogénea y que hubo interacciones múltiples entre linajes africanos y euroasiáticos, mientras que la difusión independiente de Levallois ilustraría también redes poblacionales complejas en Eurasia.
Una de las grandes implicaciones de la propuesta de Reich es que los neandertales podrían interpretarse «como humanos modernos primitivos que sufrieron una introgresión masiva de genes locales» (en palabras del autor) más que como un linaje completamente aislado. Sin entrar en debates sobre el concepto de especie, esto viene a reforzar la idea defendida por algunos de que los neandertales eran en realidad parte de Homo sapiens. En mi opinión, este escenario todavía es difícil de conciliar con el registro fósil conocido, que parece más compatible con patrones de hibridación entre dos linajes humanos próximos dentro de un proceso de especiación diferenciada a partir de un tronco común. El registro fósil neandertal no muestra morfologías claras que reflejen un origen reciente híbrido, y la morfología neandertal parece estar bien establecida desde antes de ~250 ka.
Para avanzar en este camino, futuras investigaciones deberían buscar variantes genéticas compartidas entre neandertales y humanos modernos posteriores a 300 ka, que estén potencialmente asociadas a adaptaciones cognitivas o conductuales vinculadas a tecnologías del Paleolítico medio. También debe verificarse si el reemplazo de ADNmt o cromosoma Y podría explicarse por selección natural, por ejemplo por acumulación de mutaciones perjudiciales en poblaciones neandertales pequeñas.
En cualquier caso, nuevamente aprendemos que la evolución humana en el Pleistoceno medio se caracterizó por redes complejas de interacción y de intercambio cultural, que no siempre coinciden con los episodios masivos de mezcla genética que a veces asumimos al imaginar la hibridación entre poblaciones o especies estrechamente emparentadas.
