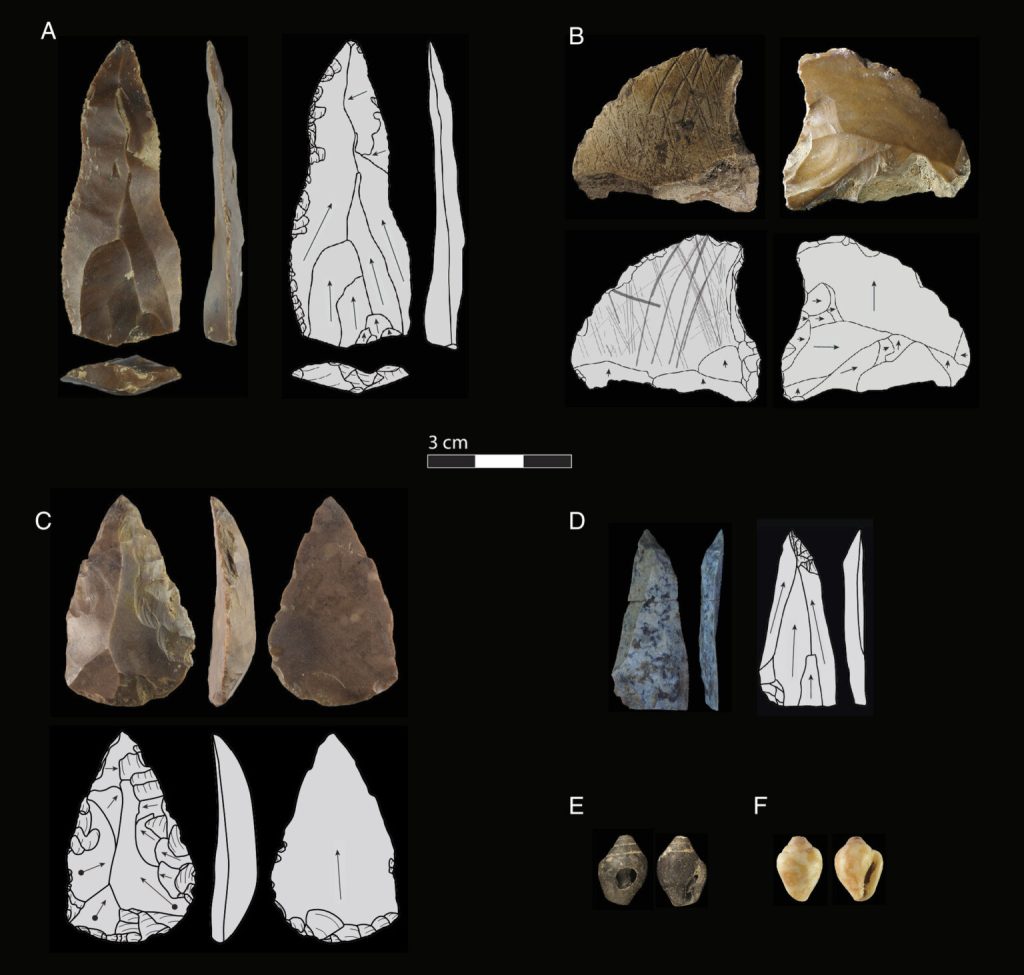
La cueva Üçağızlı II es un refugio natural en el sur de la actual Turquía. Un trabajo de Baykara y colaboradores documenta la ocupación secuencial de esta cueva por distintos grupos humanos durante unos 30.000 años: por neandertales desde 77 ka hasta 59 ka y, después, por humanos modernos desde 59 ka hasta 47 ka, sin que el registro arqueológico muestre una ruptura apreciable entre ambas ocupaciones.
Analizando más de 19.000 artefactos líticos recuperados en la cueva, los autores han encontrado que ambas especies emplearon las mismas técnicas de talla características del Paleolítico medio. Esta observación, junto con los restos de fauna y flora analizados, indica estrategias de subsistencia muy similares.
Además, se han encontrado 59 conchas de moluscos marinos (Columbella rustica) repartidas por varios niveles. Dado que este pequeño molusco carecía de relevancia por su valor alimenticio, se sugiere que las conchas se habrían seleccionado por su forma para usarlas en adornos personales. Este comportamiento no utilitario, probablemente relacionado con prácticas simbólicas, también muestra una continuidad cultural entre ambas especies.
Artefactos de la cueva de Üçağızlı II. Crédito: Baykara et al. (2026).
El año ha arrancado con un ritmo especialmente intenso en paleoantropología. A continuación resumo 49 novedades seleccionadas en este trimestre, ordenadas cronológicamente según el objeto protagonista, que ilustran las principales líneas de investigación actuales.
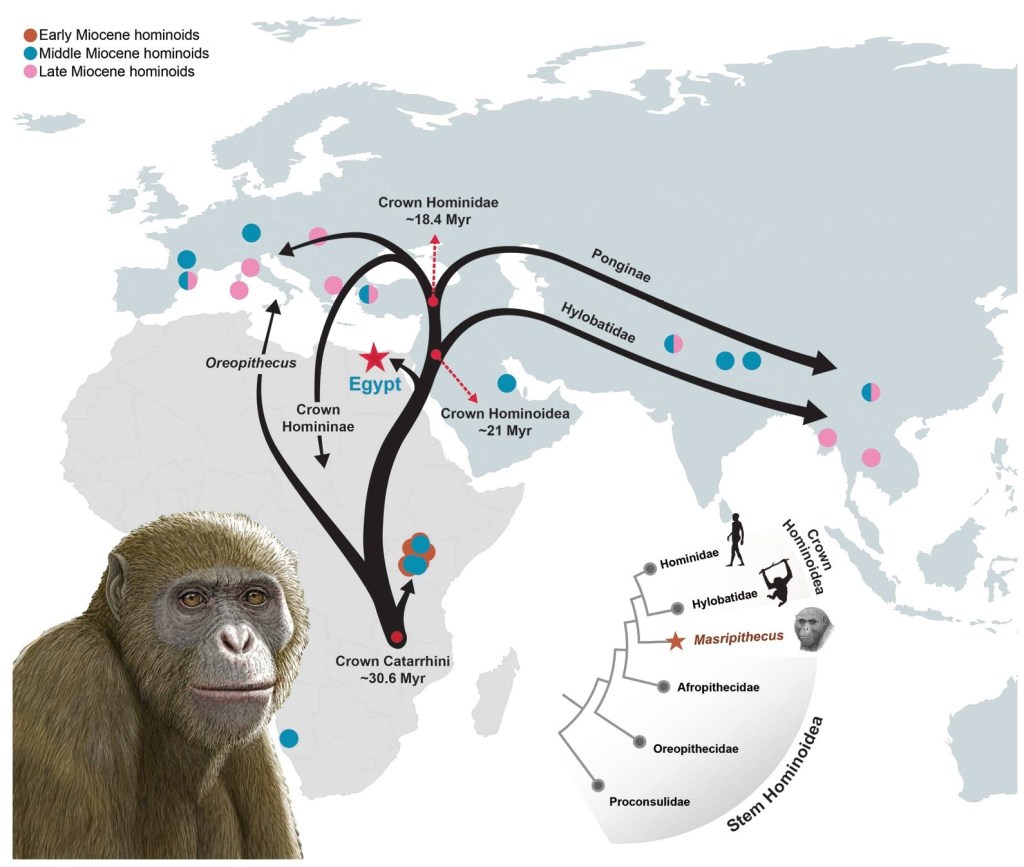
A partir de una mandíbula se ha definido una nueva especie de simio del Mioceno inicial (17-18 Ma, millones de años) en el norte de Egipto: Masripithecus moghraensis. Los autores proponen que este fósil cubre un hueco en nuestro conocimiento de la evolución de los hominoideos basales, y sitúan la parte nororiental de Afro-Arabia como posible lugar para la divergencia de este linaje antes de entrar en Eurasia, una región poco explorada en contraste con África oriental o Eurasia (Al-Ashqar et al., 2026).
Masripithecus moghraensis y la dispersión de hominoideos en el Mioceno. Crédito: M. Antón.
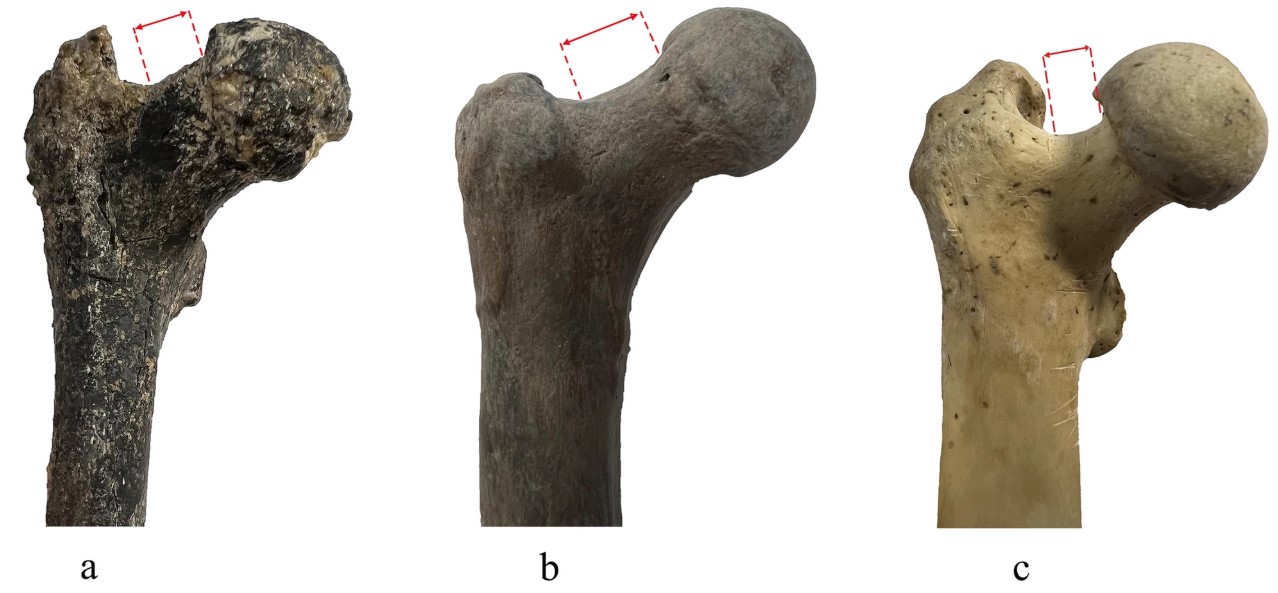
Se ha presentado un interesante fémur (Spassov et al., 2026) atribuido al género Graecopithecus, datado a finales del Mioceno (7,2 Ma) y procedente de Azmaka (sur de Bulgaría). Perteneció probablemente a una hembra de 23-24 años de edad con un peso estimado en torno a 23 kg. El fémur combina atributos propios de cuadrúpedos terrestres y de bípedos, y se agrupa principalmente con los primeros bípedos y parcialmente con los simios africanos. Entre los rasgos compatibles con una locomoción bípeda destacan el cuello femoral alargado y orientado superiormente respecto a la diáfisis, las áreas de inserción de los músculos glúteos y el grosor de la cortical ósea. Previamente, Fuss et al. (2017) ya habían identificado posibles rasgos dentales homininos en una mandíbula atribuida a Graecopithecus, como la fusión de las raíces en los premolares (no observada en los grandes simios africanos) o la reducción de los caninos. Los autores sugieren que los descendientes de este posible hominino pudieron dispersarse desde Eurasia hacia África bajo la influencia de cambios climáticos y ambientales en el Mediterráneo oriental.
Cabezas de fémur en Graecopithecus FM3549AZM6 (a), Australopithecus afarensis A.L. 288-1 (b) y chimpancé (c). Crédito: (Spassov et al. (2026).
No es novedad que el paradigma del siglo XXI en evolución humana sea un rompecabezas genético reconstruido a partir del ADN antiguo (ADNa). En el último mes, dos estudios amplían nuestro conocimiento y proponen hipótesis tentativas (siempre con la cautela de que son modelos para avanzar, no conclusiones definitivas).
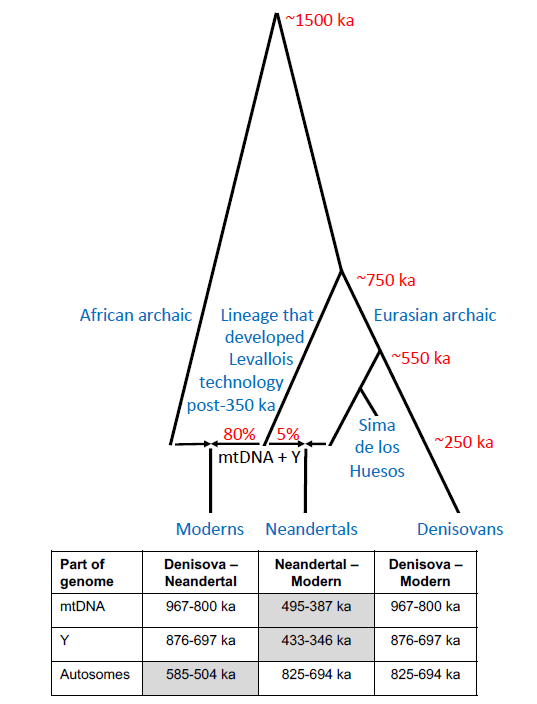
Un preprint del genetista David Reich (2026) propone un modelo demográfico para explicar aspectos de la historia de los linajes de neandertales y de humanos modernos. Según este modelo, hace entre 400 ka y 250 ka (miles de años) se produjo una expansión demográfica africana (expansión de rango o range expansion) de poblaciones asociadas a la Middle Stone Age (MSA) y la tecnología de núcleo preparado Levallois. Al expandirse hacia Eurasia, estas poblaciones habrían interactuado con grupos locales, probablemente similares a los representados en la Sima de los Huesos (Atapuerca, España), produciendo una mezcla genética muy asimétrica: la población resultante tendría aproximadamente 95% de ascendencia local basal (tipo Sima) y solo 5% de ascendencia moderna en ADN nuclear.
Aunque sería limitada esa contribución autosómica (a través de los autosomas, cromosomas no sexuales), el modelo de Reich muestra que bastaría para explicar el reemplazo completo de linajes uniparentales, como el ADN mitocondrial (ADNmt) y el cromosoma Y, debido a la dinámica de deriva y a redes de parentesco estructuradas. Las simulaciones indican que incluso una expansión numerosa puede dejar únicamente ese 5% de ascendencia autosómica africana en la población resultante. Este escenario corresponde cronológicamente a la aparición de paralelos culturales entre África y Eurasia en el Paleolítico medio, como la tecnología Levallois, el hafting (enmangado de herramientas) más sofisticado o el uso intensivo del fuego.
Hipótesis de D. Reich: Los neandertales y los denisovanos son grupos hermanos; los humanos modernos están emparentados de forma más lejana; existe una subestructura profunda en África; flujo genético hace unos 300-200 ka hacia los arcaicos euroasiáticos para dar lugar a los neandertales y, simultáneamente, hacia los arcaicos africanos para dar lugar a los modernos. Tabla: intervalos de confianza del 95% para las divergencias genéticas y las divisiones poblacionales; el resaltado en gris indica el par más cercano.
Me había propuesto en este trimestre ser realmente estricto en el número de novedades seleccionadas. Os aseguro que lo he sido, a pesar de lo cual han sido 49 las que recopilo. Estamos en un tiempo fantástico para la paleoantropología.
Las tasas de cicatrización de heridas humanas son tres veces más lentas que en primates no humanos, las cuales, a su vez, son similares a las de otras especies animales como roedores. Matsumoto-Oda y colaboradores se plantean la desventaja evolutiva que pudo haber supuesto la pérdida del vello corporal. A diferencia de los grandes simios, los humanos experimentaron en su evolución un aumento en la densidad de las glándulas sudoríparas y una disminución en la densidad del vello corporal, acompañado del engrosamiento del tejido subcutáneo como forma alternativa de protección interna. Esto habría contribuido a la evolución de tasas de cicatrización más lentas, que pueden dificultar el acceso a los alimentos, reducir la capacidad de evadir depredadores e influir sobre las necesidades de energía para la reparación de tejidos, el crecimiento y la reproducción. Es posible que el desarrollo del cuidado a individuos mayores y discapacitados entre humanos, así como el uso de plantas medicinales, mitigaran la desventaja evolutiva de la cicatrización tardía de las heridas.
Los primeros Homo (incluyendo H. habilis y H. rudolfensis) y Paranthropus boisei son propuestos como los fabricantes de útiles olduvayenses (Williams et al., 2025). Los primeros miembros del género Homo siguen siendo los principales candidatos para la fabricación y el uso del Olduvayense a lo largo de su rango temporal más amplio (c. 3,25-1,6 Ma, millones de años) pero, considerando que el Olduvayense pudo darse hasta hace 1,2 Ma, P. boisei sería el principal candidato para la producción de los artefactos posteriores.
A) Trinchera Leakey en FLK NN donde se halló el fósil OH 7 (holotipo Homo habilis). B) Núcleo olduvayense de FLK NN Nivel 3. C) OH 24, H. habilis, DK East. Crédito: CSIC-Pleistocene Archaeology Lab.
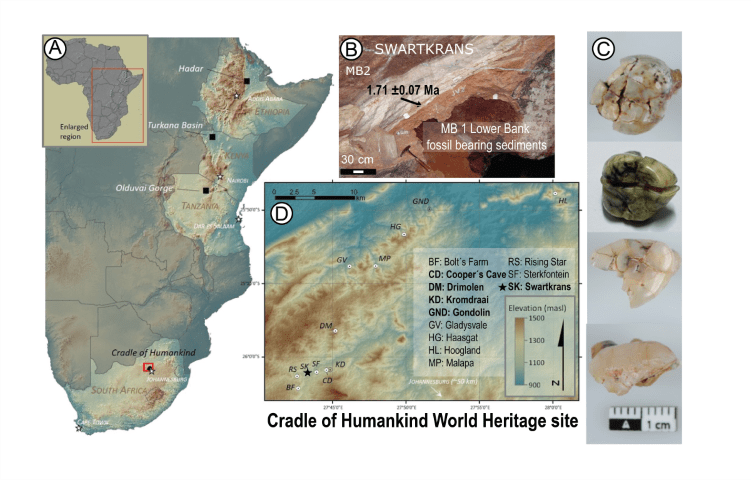
El espécimen SWT1/HR-2 de Paranthropus robustus, compuesto por un os coxae parcial articulado, el fémur casi completo y la tibia completa, procedente de Swartkrans Miembro 1 (1,7-2,3 Ma), indica bipedación completa y una estatura de solo 1,03 m, el hominino más pequeño conocido (Pickering et al., 2025).
Tras conocer el sexo biológico del espécimen Sts 63 (Australopithecus africanus) del Miembro 4 de Sterkfontein gracias a su proteoma conservado en un diente (Madupe et al., 2025), ahora han seguido el mismo camino cuatro especímenes de Paranthropus robustus de Swartkrans, de unos 2 Ma: son los dientes SK 830, SK 835, SK 850 y SK 14132. El estudio de la proteína amelogenina ha podido determinar que dos de los dientes tenían la variante específica de los machos (Madupe et al., 2025). Además, el polimorfismo de un solo aminoácido y la variación en la forma de la unión esmalte-dentina indicaron subgrupos potenciales presentes dentro de P. robustus. Anteriormente, se tenía identificada una variante genética única para la proteína polishina en dicha especie, pero este estudio mostró variaciones entre los cuatro individuos analizados. Esto podría reflejar dinámicas poblacionales complejas, microevolución o la presencia de grupos o taxones distintos.
Ubicación y estructura de Swartkrans, y vista oclusal de los dientes de Paranthropus robustus: SK 830, un P4 izquierdo; SK 835, un M3 izquierdo; SK 850, un P3 derecho; SK 14132. Crédito: Robyn Pickering.