El año ha arrancado con un ritmo especialmente intenso en paleoantropología. A continuación resumo 49 novedades seleccionadas en este trimestre, ordenadas cronológicamente según el objeto protagonista, que ilustran las principales líneas de investigación actuales.
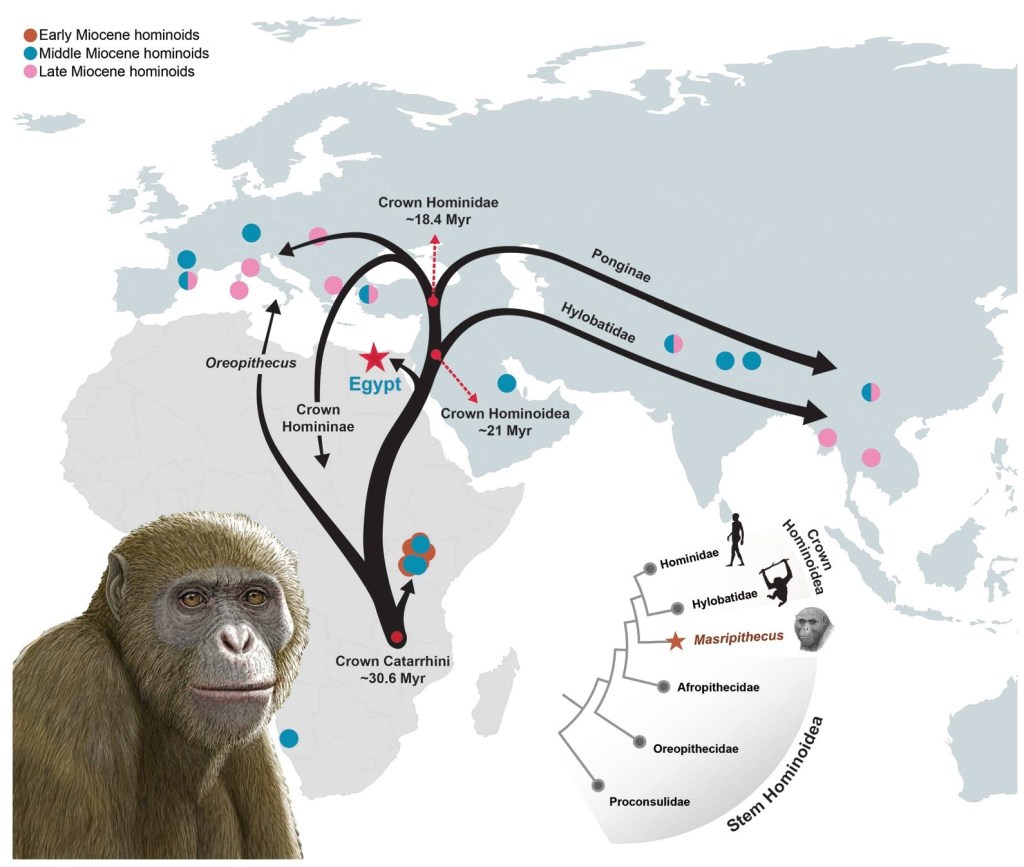
- A partir de una mandíbula se ha definido una nueva especie de simio del Mioceno inicial (17-18 Ma, millones de años) en el norte de Egipto: Masripithecus moghraensis. Los autores proponen que este fósil cubre un hueco en nuestro conocimiento de la evolución de los hominoideos basales, y sitúan la parte nororiental de Afro-Arabia como posible lugar para la divergencia de este linaje antes de entrar en Eurasia, una región poco explorada en contraste con África oriental o Eurasia (Al-Ashqar et al., 2026).
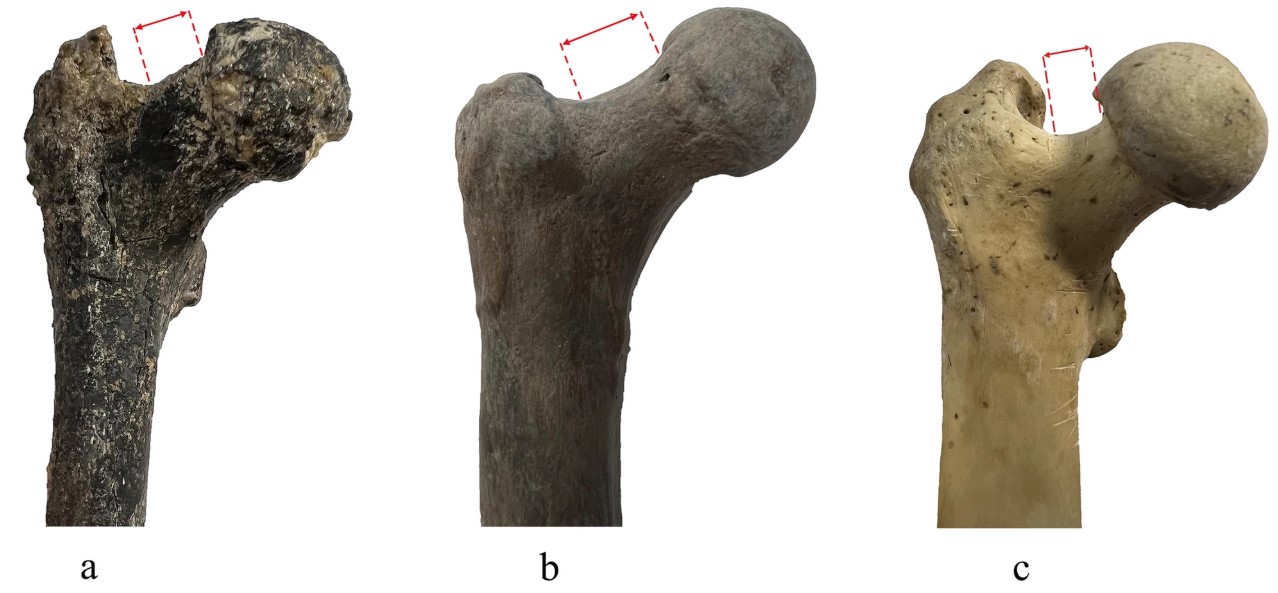
- Se ha presentado un interesante fémur (Spassov et al., 2026) atribuido al género Graecopithecus, datado a finales del Mioceno (7,2 Ma) y procedente de Azmaka (sur de Bulgaría). Perteneció probablemente a una hembra de 23-24 años de edad con un peso estimado en torno a 23 kg. El fémur combina atributos propios de cuadrúpedos terrestres y de bípedos, y se agrupa principalmente con los primeros bípedos y parcialmente con los simios africanos. Entre los rasgos compatibles con una locomoción bípeda destacan el cuello femoral alargado y orientado superiormente respecto a la diáfisis, las áreas de inserción de los músculos glúteos y el grosor de la cortical ósea. Previamente, Fuss et al. (2017) ya habían identificado posibles rasgos dentales homininos en una mandíbula atribuida a Graecopithecus, como la fusión de las raíces en los premolares (no observada en los grandes simios africanos) o la reducción de los caninos. Los autores sugieren que los descendientes de este posible hominino pudieron dispersarse desde Eurasia hacia África bajo la influencia de cambios climáticos y ambientales en el Mediterráneo oriental.