
Hasta ahora, era habitual pensar que los neandertales tardíos vivían en poblaciones pequeñas, aisladas y con claros signos de endogamia. Suelo contar esto diciendo que los Homo sapiens con quienes se iban cruzando tan solo les dieron el golpe definitivo a unas poblaciones ya debilitadas demográficamente. Esta idea se apoyaba principalmente en los genomas de varios neandertales de cuevas de los montes Altái (Siberia), como Denisova 5 (120 ka) y Chagyrskaya 8 (~80 ka), complementados por otros individuos como Okladnikov 2 (45-50 ka), Chagyrskaya 84 (~80 ka) y otros con menor cobertura. A ello se suma la hipótesis de la asimilación, según la cual algunos individuos o pequeños grupos terminaron integrándose (también genéticamente) en poblaciones sapiens.
Un nuevo trabajo de Bossoms Mesa y colaboradores matiza la propuesta del deterioro genético. El estudio abarca el ADN de 27 neandertales procedentes de diez yacimientos de Bélgica y Francia de los últimos 52,5 ka, incluyendo GN1 (Goyet), Spy94a, Z4-1514 (Les Cottés), Fond-de-Forêt 1, Trou Magrite 2423-36, entre otros. Por una parte, se han usado nuevas técnicas de descontaminación, extracción, preparación de bibliotecas genéticas y captura selectiva de fragmentos de ADN neandertal. Por otra, se ha secuenciado el quinto genoma neandertal de alta cobertura (es decir, leyendo repetidamente cada posición del ADN para minimizar errores) publicado hasta la fecha, tras los genomas de Denisova 5 (Siberia, Rusia), Vindija 33.19 (Croacia), Chagyrskaya 8 (Siberia, Rusia) y Mezmaiskaya 1 (Cáucaso, Rusia).
Uno de los indicadores genéticos más informativos se ha basado en los llamados tramos de homocigosidad o ROH (Runs of Homozygosity). Cada persona hereda una copia de cada cromosoma del padre y otra de la madre. Cuando ambos progenitores son familiares cercanos, es mucho más probable que transmitan exactamente las mismas variantes genéticas heredadas de un antepasado común. El resultado son largas regiones continuas del genoma en las que ambas copias son idénticas. Cuanto más largos y numerosos son esos tramos, más reciente e intensa ha sido la consanguinidad. Esos largos ROH en los neandertales asiáticos son los que principalmente habían conducido a la interpretación de la endogamia. Sin embargo, los neandertales tardíos del noroeste europeo no presentan apenas esos largos tramos, lo que implica que no constituían grupos pequeños aislados ni una población uniforme genéticamente, sino una metapoblación regional formada por comunidades conectadas mediante flujo génico, dentro de la cual persistieron distintos linajes durante decenas de miles de años. Estas comunidades estaban más emparentadas entre sí que con otros neandertales contemporáneos de Europa.
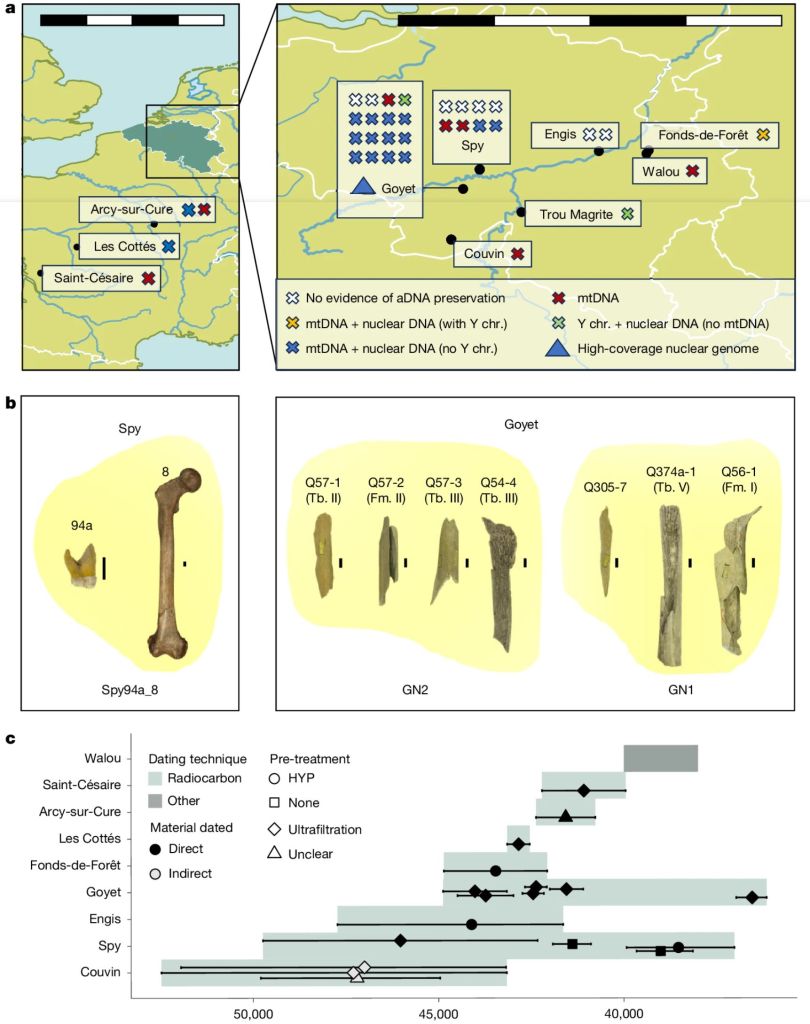
Los investigadores también analizaron el ADN mitocondrial, que solo se transmite por vía materna y permite reconstruir con buena precisión la historia de las líneas maternas. La comparación entre los distintos haplotipos reveló cambios repetidos en ellas, compatibles con movimientos de individuos entre grupos y con episodios de expansión demográfica. Además, algunos aumentos de diversidad genética coinciden con periodos climáticos más benignos, cuando las poblaciones probablemente crecían y ampliaban su área de distribución.
El trabajo confirma que los grupos de Altái eran pequeños y muy consanguíneos, escenario diferente al de los neandertales tardíos del noroeste europeo, que sí formaban una red regional de poblaciones con suficiente intercambio genético como para evitar los efectos más graves de la consanguinidad. En este sentido, los autores también estudiaron la llamada carga genética o acumulación de mutaciones perjudiciales generación tras generación, pero no detectaron un aumento progresivo de la carga genética a medida que se acercaba la extinción de la especie.
Otro resultado interesante es que algunos individuos analizados conservan fragmentos de ADN procedentes de un linaje neandertal muy antiguo, que había divergido del resto hace unos 100 ka. Es el caso de Fond-de-Forêt 1 y Trou Magrite, pero no de Goyet. Recordemos también que el neandertal Thorin de la cueva de Mandrin (Francia) representa otro antiguo linaje proveniente de aquella divergencia, que persistió hasta hace 50-42 ka, al cual también pertenece el cráneo gibraltareño de Forbes’ Quarry.
Los resultados también muestran una curiosa asimetría en la hibridación con H. sapiens. Mientras que los primeros sapiens europeos sí conservan evidencias de hibridación reciente con neandertales, los genomas de los neandertales tardíos analizados no muestran evidencias de una introgresión reciente de ADN sapiens, pese a que eran grupos contemporáneos a los primeros humanos modernos que llegaron al noroeste de Europa entre hace 45-50 ka (Ranis, Bacho Kiro, Zlatý kůň, Grotta del Cavallo, Kent’s Cavern, Oase…), con evidencias temporales más antiguas aún (Grotte Mandrin 54 ka, y las posibilidades de Bañolas 45-66 ka y de Apidima 211 ka).
En definitiva, el estudio no descarta que la genética desempeñara algún papel en la desaparición de los neandertales, pero sí debilita la hipótesis de que una decadencia genética progresiva fuera, por sí sola, la causa principal de su extinción. Los genomas siberianos dejan de considerarse el modelo de toda la especie en este aspecto. En todo caso, estas conclusiones se acotan a la muestra de individuos de Bélgica y Francia y a los últimos milenios de existencia de la especie. Es posible que en otras regiones existieran poblaciones mucho más pequeñas y aisladas, con endogamia predominante, como por ejemplo parecen indicar los neandertales de El Sidrón, en el norte de España, de hace unos 49 ka (Lalueza-Fox et al., 2010).
