El reciente estudio de Jakobsson y colaboradores sobre 28 genomas antiguos de individuos procedentes de distintas regiones del sur de África, permite observar una historia poblacional que permaneció sorprendentemente estable durante el Holoceno. La muestra abarca casi nueve milenios (entre hace ~10 ka y ~1 ka), a través de los cuales existe una continuidad genética temporal, muy baja estratificación espacial, pero también diversidad interna.
Las divergencias poblacionales promedio entre individuos que representan a este grupo meridional antiguo con cualquier otro grupo (africanos occidentales, orientales, centrales, septentrionales y no africanos antiguos y modernos) se estiman en alrededor de 310-240 ka. Además, la afinidad genética entre estos meridionales antiguos y los africanos orientales (antiguos y actuales) es similar a su afinidad genética con los occidentales (antiguos y actuales). Esto hace poco probable un flujo genético detectable de cualquiera de estos grupos desde alrededor de 150-200 ka, periodo en que suele identificarse la divergencia entre los africanos occidentales y orientales.
Por tanto, los cazadores-recolectores de África meridional, especialmente los anteriores a 1,4 ka, habrían permanecido con un significativo aislamiento genético durante al menos 200 ka. Esto implica que aquella región habría sido refugio geográfico duradero para una gran población humana hasta la llegada, hace unos 2 ka, de agricultores y pastores neolíticos procedentes del norte, documentados tanto genética como arqueológicamente, junto con grupos occidentales y, más tarde, poblaciones con ascendencia europea. Este linaje meridional es uno de los más divergentes conocidos dentro de nuestra especie, incluyendo los grupos Khoe-San actuales.
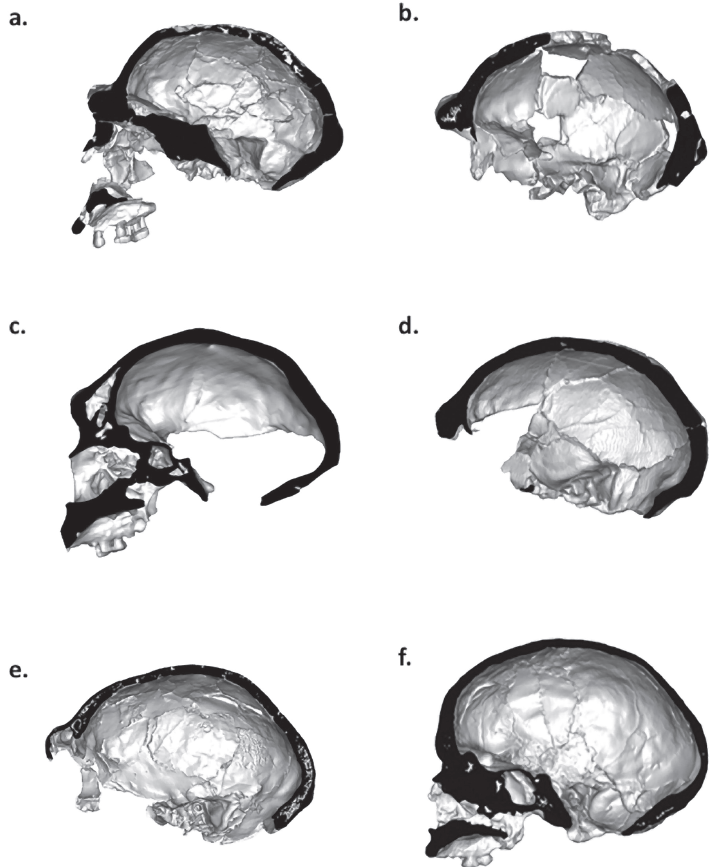
Una de las características más llamativas de Homo sapiens es nuestra frente alta y vertical. La combinación de una serie de rasgos nos distingue frente a otras especies humanas, tales como el cráneo globular, la mencionada frente, el arco superciliar reducido o ausente, la cara retraída, la fosa canina y el mentón pronunciado.
El hueso frontal refleja patrones evolutivos de las poblaciones del Pleistoceno, de forma que H. erectus y H. neanderthalensis tienen frentes inclinadas y toros supraorbitales prominentes, mientras que H. sapiens presenta una frente más vertical y una reducción significativa del torus. Curiosamente, las variaciones suelen ser más evidentes en los laterales del hueso frontal que en su región media.
La evolución de nuestro frontal vertical se ha relacionado con distintos factores, como una combinación de cambios en la base craneal, el crecimiento cerebral y el desarrollo posnatal en nuestra especie.
Cambios en el esfenoides y en la flexión de la base craneal
El esfenoides es el hueso central de la base del cráneo, a partir del cual crece la cara hacia adelante. El acortamiento del esfenoides en nuestra ontogenia, en comparación con el de otros homininos, fue visto desde hace tres décadas como causa de la retracción facial y de una configuración craneal más globular, con un aumento del ángulo entre la base craneal y la parte superior del cráneo. Sin embargo, aunque el acortamiento del esfenoides contribuye al cambio, es insuficiente por sí solo para explicar la totalidad de la transformación hacia una frente alta.
La flexión de la base craneal consiste en el proceso de curvatura que se produce particularmente en la interacción entre el esfenoides y el occipital. La sincondrosis esfeno-occipital es un punto de crecimiento cartilaginoso entre estos huesos, eje sobre el que descansa la biomecánica del cráneo, y juega un papel clave en esta reorganización. En H. sapiens, la prolongación del crecimiento de esta estructura durante la ontogenia permite una mayor adaptación a las demandas del crecimiento cerebral.
Una mayor flexión reduce la longitud de la base anterior, eleva la frente y facilita una reorganización más uniforme del volumen craneal, para alojar un cerebro más globular. El aumento relativo del volumen de los lóbulos temporales también podría estar relacionado con esta flexión, y tener un posible vínculo con funciones cognitivas y lingüísticas asociadas a estas regiones.
Desarrollo posnatal
Se ha observado en distintas especies una correlación entre las vías que sigue el desarrollo posnatal y la altura del neurocráneo. Comparado con sus antepasados, el género Homo experimentó una trayectoria única de desarrollo en la que retiene características juveniles en la etapa adulta, como proporciones neurocraneales elevadas. Especies como H. naledi y H. longi muestran un crecimiento negativo del neurocráneo respecto a la cara, un rasgo arcaico. Este punto de partida ontogenético cambia en H. sapiens. Es evidente que los neandertales y los humanos modernos alcanzan tamaños cerebrales adultos comparables a través de distintas trayectorias de desarrollo, en las evoluciones paralelas de estos linajes «primos».
La expansión del cerebro durante los primeros años contribuye a reorganizar la estructura craneal y al desplazamiento vertical de la frente. Por ejemplo, las fontanelas son espacios abiertos entre los huesos del cráneo que permanecen sin osificar durante los primeros años de vida. En H. sapiens, las fontanelas se mantienen abiertas por más tiempo en comparación con otras especies humanas. Este fenómeno facilita el crecimiento cerebral postnatal y permite un remodelado craneal que favorece la formación de una frente alta.
Variación del hueso frontal en distintas especies humanas: a) KNM-ER 3733 (H. erectus); b) Ceprano (H. neanderthalensis); c) Broken Hill (H. rhodesiensis); d) Laetoli 18 (basal? H. sapiens); e) Spy 1 (H. neanderthalensis); f) Mladeč 1 (H. sapiens). Crédito: Athreya (2012).
Reorganización cortical y crecimiento cerebral
El tamaño del cerebro de H. sapiens también importa. Con un volumen cerebral promedio de unos 1350 cc, nuestra especie supera en capacidad a la mayoría de los homininos arcaicos, salvo a H. neanderthalensis y a H. juluensis (denominación que viene a significar «cabeza enorme»). El cráneo juvenil de H. sapiens habría sido un motor de este mayor volumen cerebral relativo y de la frente más vertical, sin requerir un aumento absoluto de la masa encefálica.
En particular, la expansión de los lóbulos frontales genera una mayor demanda de espacio en la región anterior del cráneo. El lóbulo frontal no solo ha aumentado en volumen, sino que también ha sufrido una reorganización funcional, que incluye la elevación de la frente. Por ejemplo, la reconstrucción digital de los cráneos de Buia y Bodo muestra que la forma del frontal no depende exclusivamente del tamaño cerebral, sino de restricciones mecánicas y topográficas del cráneo. Esto afecta al desarrollo de áreas asociadas con la cognición avanzada en la corteza prefrontal.
Factores biomecánicos y tensión funcional
Finalmente, se ha propuesto que la disminución de las tensiones mecánicas en el cráneo también ha podido influir en la morfología de la frente. Las fuerzas generadas durante la masticación crean una presión que puede estar relacionado con el prognatismo facial marcado y la prominencia del torus supraorbital. En H. sapiens, la reducción del prognatismo podría haber reducido parte de estas tensiones, favoreciendo una frente más elevada. Sin embargo, también se ha relacionado el desarrollo del torus supraorbital con cambios en la relación espacial entre el neurocráneo y la cara, más que a tensiones funcionales masticatorias o cargas musculares.
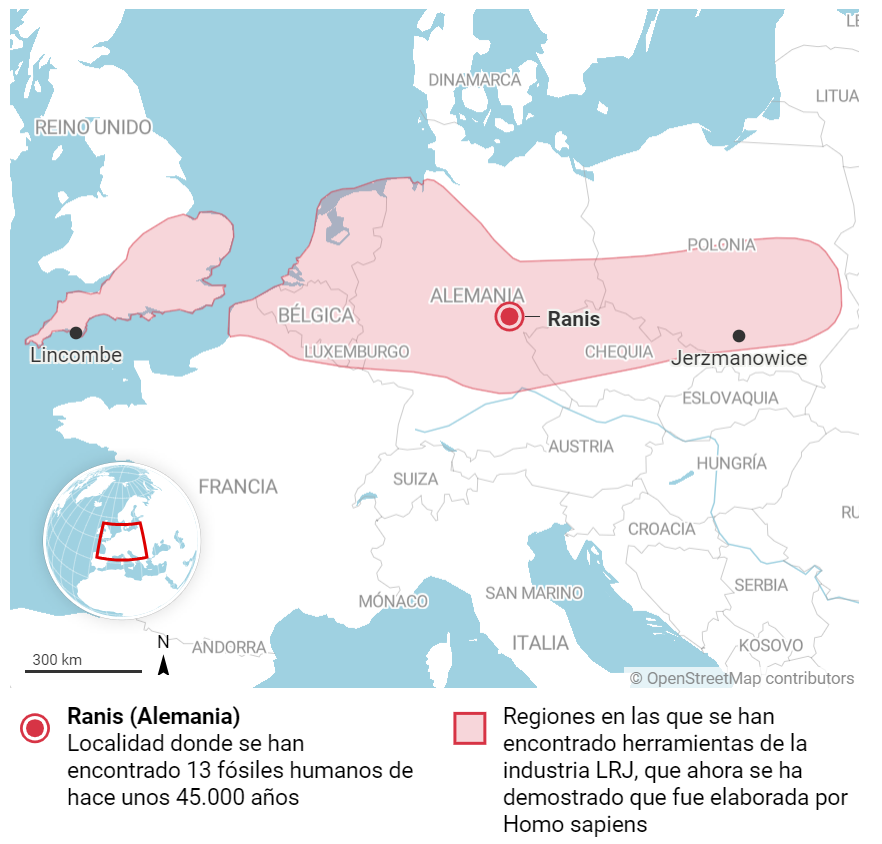
Entre 1932 y 1938, en el yacimiento de Ilsenhöhle, en Ranis (Turingia, Alemania) se hallaron herramientas de piedra vinculadas a un periodo cultural de transición entre el Paleolítico medio y el Paleolítico superior conocido como Lincombiano-Ranisiano-Jerzmanowskiano (LRJ). Este tecnocomplejo se ha encontrado también por otros sitios del centro y noroeste de Europa, con distintas dataciones entre 44 y 41 ka. Aunque esta cultura se venía asociando (con dificultades) a la especie Homo neanderthalensis, basándose en su antigüedad, un nuevo trabajo ha resuelto muchas dudas a partir de los resultados de una nueva excavación en Ilsenhöhle que tuvo lugar entre 2016 y 2022.
Localización del yacimiento de Ilsenhöhle y área de hallazgo del tecnocomplejo LRJ: Crédito: La Vanguardia & Mylopotamitaki, D., Weiss, M., Fewlass, H. et al. (2024). Homo sapiens reached the higher latitudes of Europe by 45,000 years ago. Nature.
Mediante tres publicaciones, se analizan nuevas herramientas del LRJ, junto con restos óseos de reno, bisonte, rinoceronte lanudo, oso de las cavernas, hiena, glotón, zorro ártico, lobo y glotón, y un total de 13 fragmentos humanos, incluidos algunos de la excavación 1932-38. A partir del ADN mitocondrial recuperado de 11 fragmentos (7 de la excavación 1932-38 y 4 de la 2016-22), se ha conseguido determinar que estos restos son de H. sapiens. La datación molecular del material genético resulta entre 49-41 ka, consistente con la datación por radiocarbono de 47-42 ka de las capas sedimentarias de donde proceden. El material genético también ha permitido relacionar a este grupo con el individuo sapiens de Zlatý kůň (República Checa), de unos 45 ka, que por cierto tiene un 3% de rastro neandertal en su genoma (no se reporta rastro neandertal en los restos de Ranis). Sigue leyendo →
Hasta ahora se conocen cinco especies humanas que han habitado los distintos sitios de Atapuerca a lo largo de casi un millón y medio de años. Descúbrelas en esta conferencia impartida para el Club de Ciencia Boadilla el 29 de septiembre de 2023:
Aunque distintos estudios van evidenciando un origen africano de Homo sapiens, no está nada claro cómo y cuándo sucedió, y cómo fue la posterior evolución de nuestra especie. Según vamos teniendo más genomas secuenciados, vemos menos clara la posibilidad de una divergencia relativamente reciente a partir de una sola población ancestral en África, ya sea en el este o en el sur, como nos parecía hace 5-10 años. Además, es difícil conciliar la visión genética con los registros fósiles y arqueológicos de ocupaciones humanas antiguas identificadas por todo el continente africano. Unos pocos ejemplos que ilustran ese problema son los cráneos de Jebel Irhoud en el norte (~300 ka, miles de años), Omo (~230 ka) y Herto (~165 ka) en el este, y Florisbad (~259 ka) en el sur. Desde hace más de 300 ka, las características anatómicas derivadas de Homo sapiens se van encontrando de forma dispersa y desigual en todo el continente. Trabajos como el de Mounier y Mirazón Lahr (2019) tratan de modelizar distintas fases de la evolución de nuestro linaje a partir de la morfología de los cráneos arcaicos y modernos, como los mencionados anteriormente, y evidencian la dificultad para concretar resultados dado lo reducido del tamaño del registro fósil. Y está por determinar el encaje en todo esto de otra especie que genera muchos interrogantes, Homo naledi (Sudáfrica, 241-335 ka), así como de especímenes más alejados de rasgos sapiens, como el cráneo Kabwe-1, asociado a Homo rhodesiensis.
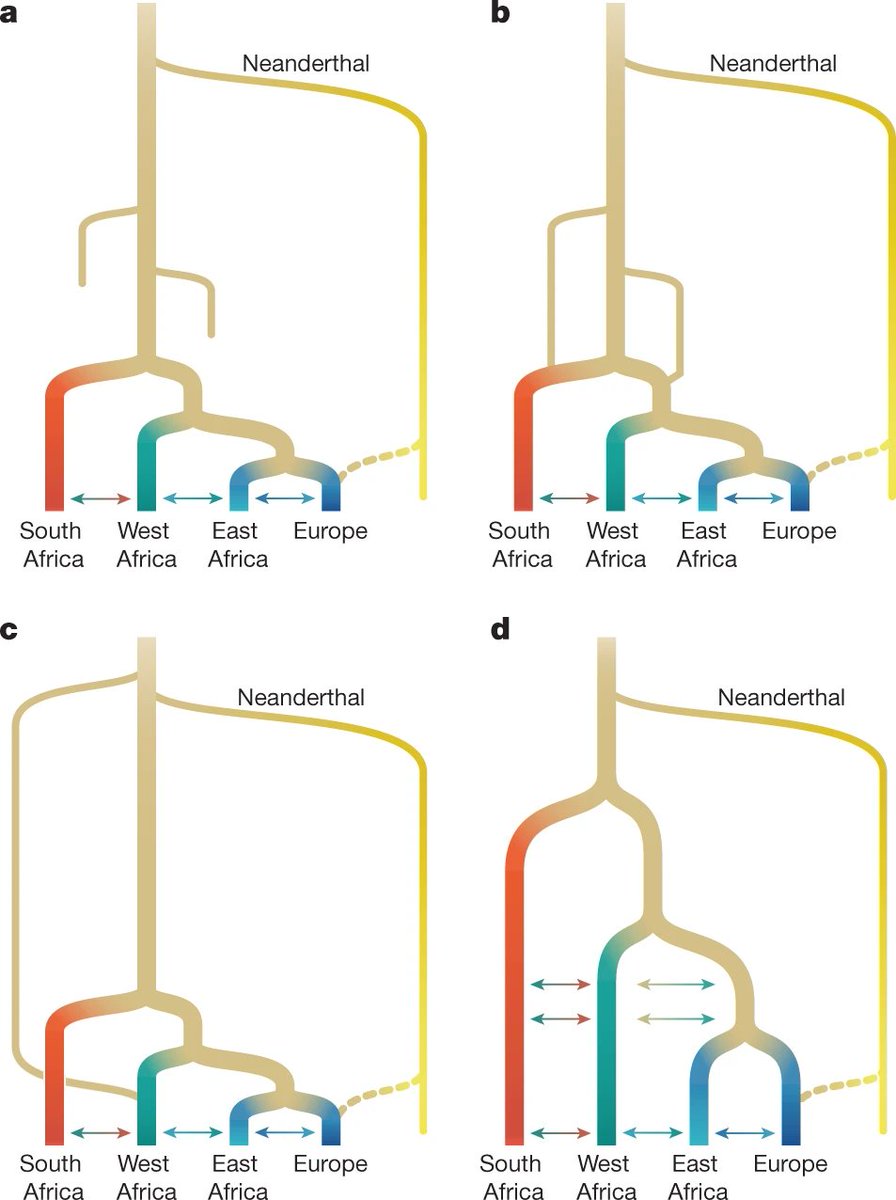
Los modelos de la literatura paleoantropológica traducibles en modelos demográficos genéticamente comprobables, se pueden ordenar en cuatro grandes posibilidades: a) una expansión reciente; b) una expansión reciente con persistencia regional; c) una mezcla arcaica; d) un desarrollo multirregional africano:
Crédito: Ragsdale, A.P., Weaver, T.D., Atkinson, E.G. et al. (2023)
A partir de un algoritmo inicialmente diseñado para tratar de entender el riesgo genético de enfermedad entre distintas poblaciones, los autores de un nuevo trabajo (A. P. Ragsdale, T. D. Weaver, E. G. Atkinson et al., 2023) reorientaron su utilización para profundizar en los orígenes de los humanos modernos. Con él, probaron cientos de escenarios posibles de flujo genético entre poblaciones en distintas partes de África. Como datos, se emplearon los genomas de 290 individuos de cuatro grupos actuales: 44 nuevos genomas secuenciados de los nama (khoe-san de Sudáfrica), 85 de los mende (de Sierra Leona), 23 de los gumuz (descendientes recientes de un grupo de cazadores-recolectores de Etiopía) y 46 de agricultores de África oriental (amhara y oromo de Etiopía, muy similares genéticamente). También se incluyeron 91 individuos británicos, como fuente representativa del flujo genético de vuelta a África y la mezcla colonial reciente en Sudáfrica, y un genoma neandertal de la cueva de Vindija (Croacia). Las conclusiones se pueden resumir así: Sigue leyendo →