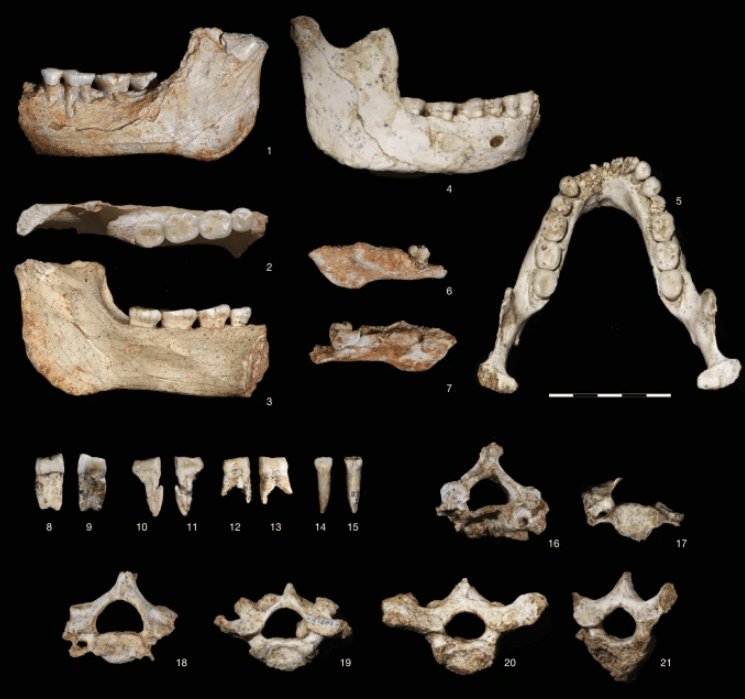
A partir de cinco fragmentos craneales/faciales y cinco dientes, se ha logrado reconstruir virtualmente el cráneo DAN5/P1 (Baab, Kaifu, Freidline et al., 2025). El fósil procede de Gona (Etiopía) y está datado en 1,6-1,5 Ma. Este trabajo ha dado como resultado uno de los cráneos de Homo más completos de su marco temporal.
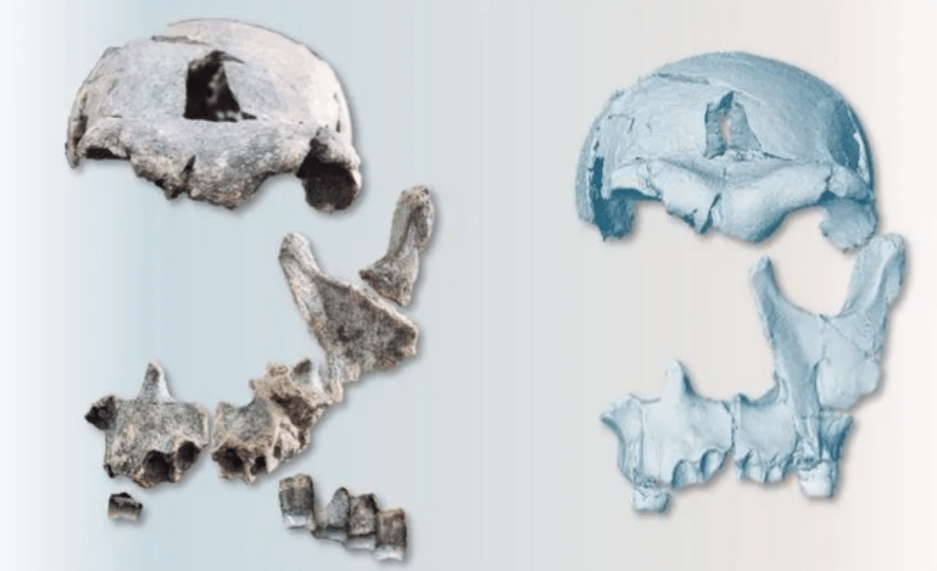
DAN5 presenta un mosaico morfológico que, una vez más, cuestiona visiones evolutivas lineales entre Homo habilis y Homo erectus. Sus principales características son:
- Cráneo pequeño, con un volumen de unos 598 cc, claramente por debajo del promedio de H. erectus africano clásico, solapándose con el de H. habilis y otros Homo tempranos.
- Rasgos típicos de H. erectus, como un toro supraorbital desarrollado, perfil craneal bajo y alargado, glabela marcada y parietales relativamente rectos.
- Cara corta y relativamente ortognata, con plano medio casi vertical, pómulos transversalmente planos y menor prognatismo que en muchos Homo tempranos, aunque sin los rasgos derivados de H. sapiens.
- Región nasal plana y estrecha, similar a la de H. habilis. La baja posición de la nariz y de la cara en general recuerda a OH 24 (H. habilis) y D4500 (H. erectus Dmanisi), en contraste con las caras más altas y robustas de KNM-ER 3733 y ER 3883 (H. erectus africanos).
- Paladar anterior ancho y casi cuadrangular, con arcada dental relativamente expandida anteriormente, consistente con condiciones plesiomorfas dentro de Homo.
- Dentición y mandíbula con proporciones y robustez cercanas a Homo temprano. El tamaño relativo de molares y premolares es una plesiomorfía respecto a H. erectus africanos.