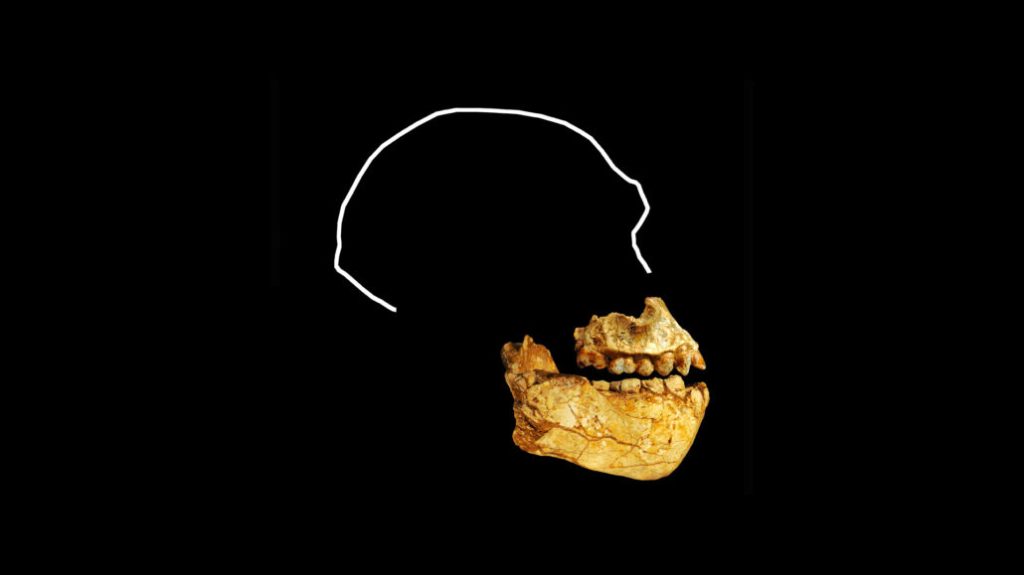
El reciente estudio de Jakobsson y colaboradores sobre 28 genomas antiguos de individuos procedentes de distintas regiones del sur de África, permite observar una historia poblacional que permaneció sorprendentemente estable durante el Holoceno. La muestra abarca casi nueve milenios (entre hace ~10 ka y ~1 ka), a través de los cuales existe una continuidad genética temporal, muy baja estratificación espacial, pero también diversidad interna.
Las divergencias poblacionales promedio entre individuos que representan a este grupo meridional antiguo con cualquier otro grupo (africanos occidentales, orientales, centrales, septentrionales y no africanos antiguos y modernos) se estiman en alrededor de 310-240 ka. Además, la afinidad genética entre estos meridionales antiguos y los africanos orientales (antiguos y actuales) es similar a su afinidad genética con los occidentales (antiguos y actuales). Esto hace poco probable un flujo genético detectable de cualquiera de estos grupos desde alrededor de 150-200 ka, periodo en que suele identificarse la divergencia entre los africanos occidentales y orientales.
Por tanto, los cazadores-recolectores de África meridional, especialmente los anteriores a 1,4 ka, habrían permanecido con un significativo aislamiento genético durante al menos 200 ka. Esto implica que aquella región habría sido refugio geográfico duradero para una gran población humana hasta la llegada, hace unos 2 ka, de agricultores y pastores neolíticos procedentes del norte, documentados tanto genética como arqueológicamente, junto con grupos occidentales y, más tarde, poblaciones con ascendencia europea. Este linaje meridional es uno de los más divergentes conocidos dentro de nuestra especie, incluyendo los grupos Khoe-San actuales.
Sigue leyendo