
Este trimestre que cerramos ha venido cargado de novedades: nada menos que 49 trabajos recopilados, que muestran cómo este campo sigue despertando interés, tanto en la comunidad científica como en el público general. Ahora bien, siguiendo mi campaña contra los titulares exagerados, conviene recordar que cada hallazgo no supone “reescribir la historia de la evolución humana”. ¿Lo habríamos tenido que hacer 49 veces en un solo trimestre? Comprendamos el significado de cada avance en su contexto.
- El ilion experimentó dos innovaciones clave en la evolución humana que hicieron posible la bipedación: la reorientación de la placa de crecimiento ilíaca perpendicular a la columna vertebral, y una osificación ilíaca humana tardía (Senevirathne, Fernandopulle, Richard et al., 2025).
- Los chimpancés del valle de Issa (Tanzania) pasan tanto tiempo en los árboles como los chimpancés de bosques más densos. Además, no solo trepan, sino que también suelen ponerse de pie y desplazarse erguidos sobre las ramas, usando los brazos para equilibrarse mientras buscan alimento. Se trata de un entorno parecido al de los primeros homininos de hace entre 4 y 7 Ma (millones de años), que aún mostraban adaptaciones claras para trepar, como brazos largos y dedos curvados. Aquellos homininos pudieron haber combinado vida arbórea y terrestre y desarrollado el caminar erguido en las ramas, aprendiendo el equilibrio antes de dominar la marcha en espacios abiertos (Drummond-Clarke et al., 2025).
- Baker, Barton y Venditti han hallado en 95 primates fósiles y actuales una correlación entre la longitud del pulgar, una característica clave del agarre de precisión, y el tamaño del cerebro, utilizando métodos filogenéticos bayesianos. En concreto, no se ha encontrado un vínculo con el tamaño del cerebelo, pero sí una fuerte relación con el tamaño del neocórtex, lo que puede reflejar el papel de las cortezas motora y parietal en las habilidades sensomotoras asociadas con la manipulación fina.
- El análisis de ocho dimensiones poscraneales sugiere que tanto Australopithecus afarensis como Australopithecus africanus presentan un alto grado de dimorfismo sexual de tamaño, significativamente mayor que el de los chimpancés y los humanos modernos. A su vez, Au. afarensis tiene mayor dimorfismo que Au. africanus (Gordon, 2025).
- El estudio de las clavículas de Au. afarensis, en particular las del individuo juvenil DIK-1-1 y del adulto KSD-VP-1/1, en comparación con las de simios actuales, revela una geometría cortical que sugiere un comportamiento arbóreo sostenido en estos homininos, incluyendo suspensión y escalada (Farrell y Alemseged, 2025).
- La introducción en la dieta de los australopitecos de vegetales duros como las gramíneas precedió en unos 700 ka a la adaptación de su dentición, con el desarrollo de molares más largos y la reducción del tamaño de los dientes en un 5% cada 1000 años. Fannin y colaboradores han observado que los Au. afarensis dejaron de consumir frutas, flores e insectos para alimentarse de pastos y juncos hace entre 3,4 y 4,8 Ma, a pesar de carecer de los dientes y el sistema digestivo óptimos para comer estas plantas más duras y resistentes. Los homininos habrían podido maximizar los nutrientes derivados de estas plantas a medida que sus dientes se volvían más eficientes para descomponerlas. Posteriormente, hace unos 2 Ma los isótopos de carbono y oxígeno en los dientes de los primeros Homo cambiaron abruptamente, por la reducción del consumo de pastos y el aumento relativo del agua probablemente contenida en tubérculos, bulbos y cormos, tejidos vegetales de alta energía. En este momento los dientes muestran un cambio repentino en forma y tamaño más adecuado para comer tejidos cocinados.
- Los homininos de Nyayanga (Kenia) fabricaron herramientas olduvayenses hace 2,6 Ma principalmente a partir de materia prima no local, transportada hasta 13 km para aprovechar su calidad (Finestone et al., 2025). Con esta antigüedad, los autores de aquellas herramientas pudieron ser representantes de Homo tempranos o Paranthropus, cuya presencia se ha documentado en el mismo nivel NY-1. Las anteriores evidencias de transporte de materias primas estaban datadas en unos 2 Ma, procedentes de Kanjera South (a 15 km de Nyayanga) y de Olduvai y Ewass Oldupa en Tanzania. El mismo trabajo recopila una comparativa de lugares y distancias de transporte. Se han observado transportes similares realizados por chimpancés durante unos 2 km.

- El análisis de marcas sobre dos fósiles de Homo habilis (OH 7 y OH 65) con el apoyo de modelos de IA entrenados con una biblioteca de 1496 imágenes de marcas dentales de carnívoros modernos (leopardos, leones, cocodrilos, lobos, hienas) indica que los leopardos eran los responsables de las marcas de los dientes, con un 90 % de probabilidad. Por tanto, la posición trófica de algunos de los primeros representantes del género Homo no difería de la de otros australopitecos ya que, si H. habilis se hubiera convertido en una especie competidora con los grandes carnívoros, sus huesos probablemente habrían sido devorados por animales trituradores de huesos, como las hienas, después de que murieran por otras causas. Sin embargo, los leopardos los cazaban activamente, lo que sugiere que la transición a una posición dominante en la cadena alimentaria se produjo posteriormente (Vegara‐Riquelme, Baquedano y Domínguez-Rodrigo, 2025).
- En 2022 se había anunciado en prensa el hallazgo de un molar en Qvemo Orozmani, Georgia, muy cercano a Dmanisi y de una antigüedad similar (1,77-1,84 Ma). Ahora se anuncia el descubrimiento de una mandíbula en el mismo yacimiento. Esperemos la publicación de estos fósiles.
- Se han documentado siete artefactos líticos procedentes de Calio (Sulawesi) que sitúan a grupos humanos en Wallacea de forma más amplia y anterior a lo que se sabía (Hakim, Wibowo, van den Bergh et al., 2025). En 2016 ya se habían documentado artefactos líticos en Talepu de al menos 118 ka, dejando claro que Sulawesi, al igual que Flores y Luzón, había estado ocupada por homininos mucho antes de la llegada de los humanos modernos a la región. Los nuevos artefactos son lascas simples, algunas retocadas, y están datados en 1,04-1,48 Ma. Dado que el canal de aguas profundas llamado Línea de Wallace siempre ha mantenido separadas Sulawesi, Lombok y Flores del resto de masa continental cuando el mar fluctuaba 130 metros por debajo del nivel actual durante el Pleistoceno, esto implica que los humanos llegaron a Sulawesi hace al menos un millón de años.

- Engaji Nanyori (Lecho III, Garganta de Olduvai, Tanzania) ofrece una visión poco común del Achelense durante la transición al Pleistoceno medio (1,1-0,9 Ma), un periodo de cambio climático y aridificación en el que estos hallazgos son raros. En este momento, los Homo erectus realizaban producción estable de lascas con estrategias de talla similares al Olduvayense, LCT poco frecuentes y una sorprendente uniformidad panafricana (Cueva-Temprana et al., 2025).
- La reconstrucción y análisis del cráneo Yunxian 2, fechado en 1 Ma, le posiciona como un posible representante temprano del clado longi (Peng et al., 2025). Su mezcla de rasgos podría estar indicando que era cercano al ancestro común de las líneas evolutivas longi–sapiens (rasgos apomórficos) y erectus-heidelbergensis (plesiomorfias).

- Hace 430 ka (miles de años), poblaciones humanas en Marathousa 1 (Grecia) tallaron microlitos, pequeñas herramientas versátiles, tanto lascas simples como retocadas, adaptadas al uso específico de procesamiento de restos del elefante Palaeoloxodon antiquus, y elaboradas con talla a mano alzada y bipolar (De Caro et al., 2025).
- Los restos faunísticos del nivel TD10.2‑BB de Gran Dolina (Atapuerca, España) indican que, hace más de 400 ka, grupos humanos organizaban cacerías comunales de bisontes de gran envergadura. Se han identificado al menos 60 individuos abatidos sin discriminación por edad, cazados en periodos clave del año (finales de primavera y principios de otoño) cuando el rendimiento energético por animal era óptimo. Estas matanzas podrían abastecer a grupos de hasta 30 individuos durante semanas, con una eficiencia difícilmente alcanzable sin una cooperación estructurada y compleja. A pesar de la intensidad de estas prácticas, las curvas demográficas de los bisontes no reflejan signos de sobreexplotación, sino una dinámica poblacional saludable, lo que sugiere una forma de aprovechamiento ecológicamente sostenible, aunque no planificada como sostenible en el sentido moderno del término (Rodríguez-Gómez, Rodríguez-Hidalgo, Saladié et al., 2025).
- El estudio de los 21 dientes de Hualongdong (Wu, Martinón-Torres, Xing et al., 2025) indica que la mayoría de sus rasgos no se parecen a los neandertales y sí a los de homininos del Pleistoceno final, como el tercer molar reducido, a pesar de su edad del Pleistoceno medio. Se exceptúan las raíces robustas de los premolares y molares, que sí se aproximan a las morfologías de su periodo. En general, estas características no están presentes en la mayoría de especímenes del Pleistoceno medio tardío en Asia oriental (300-120 ka), salvo quizás en los de Panxian Dadong o Jinniushan.
- Se ha presentado un conjunto de 35 herramientas de madera de 300 ka halladas junto al lago Fuxian en Gantangqing, al suroeste de China (Liu et al., 2025). Se trata de palos y otras herramientas con punta para excavar raíces, extraer tubérculos y recolectar alimentos vegetales del subsuelo. Se encontraron en asociación con restos vegetales, como piñones, avellanas, frutas y rizomas acuáticos, lo que sugiere una actividad de recolección repetida y organizada. Recordemos que entre las herramientas contemporáneas de Schöningen (Alemania), aproximadamente la mitad serían usadas para cazar grandes animales, pero hay otro buen número hechas con maderas hendidas y terminación puntiaguda o redondeada, probablemente empleadas también en actividades domésticas de procesamiento de vegetales y pieles.

- El pequeño tamaño de Homo floresiensis pudo haber evolucionado ralentizando su crecimiento durante la infancia, no en el desarrollo prenatal (Kaifu et al., 2025). El tamaño del tercer molar es pequeño en proporción a los demás molares, lo cual es típico en el género Homo. El desarrollo fetal parece que estaría encaminado a desarrollar los cerebros grandes que caracterizan a Homo, y la ralentización del crecimiento cerebral en H. floresiensis probablemente ocurrió después del nacimiento.
- Se ha datado en al menos 286 ka el cráneo de Petralona, por el método de series de uranio sobre la calcita que creció directamente sobre el cráneo (Falguères et al., 2025).
- Atendiendo a la evolución de la tecnología Levallois en los yacimientos de Nor Geghi-1 (Armenia) y Purfleet (Reino Unido), la aparición de esta tecnología parece resultado de procesos distintos en ambas regiones, respaldando un modelo de origen múltiple en los sistemas de producción de núcleos preparados en el Paleolítico inferior (Gill et al., 2025).
- Una herramienta ósea fue elaborada por neandertales a partir de un hueso de león de las cavernas (Panthera spelaea) hace más de 130 ka en la cueva de Scladina (Bélgica), como parte de un conjunto complejo de útiles que integraba tecnologías líticas y óseas. Se trata de una tibia con marcas de modificación intencionada asociadas a una doble secuencia de uso: primero como instrumento de raspado o procesamiento y posteriormente como retocador de herramientas líticas. El uso técnico de restos de grandes carnívoros por parte de neandertales es una práctica extremadamente rara en el registro arqueológico (Abrams, Auguste, Pirson et al., 2025).

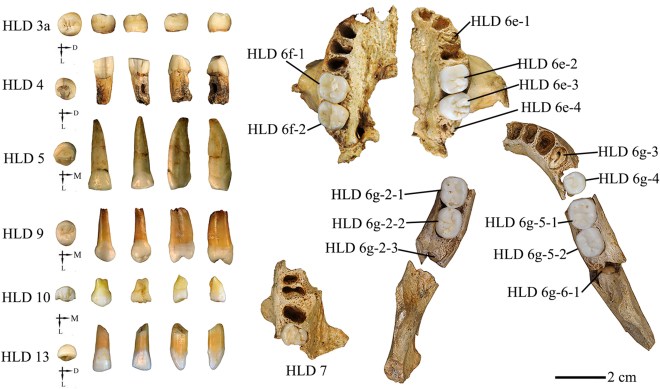
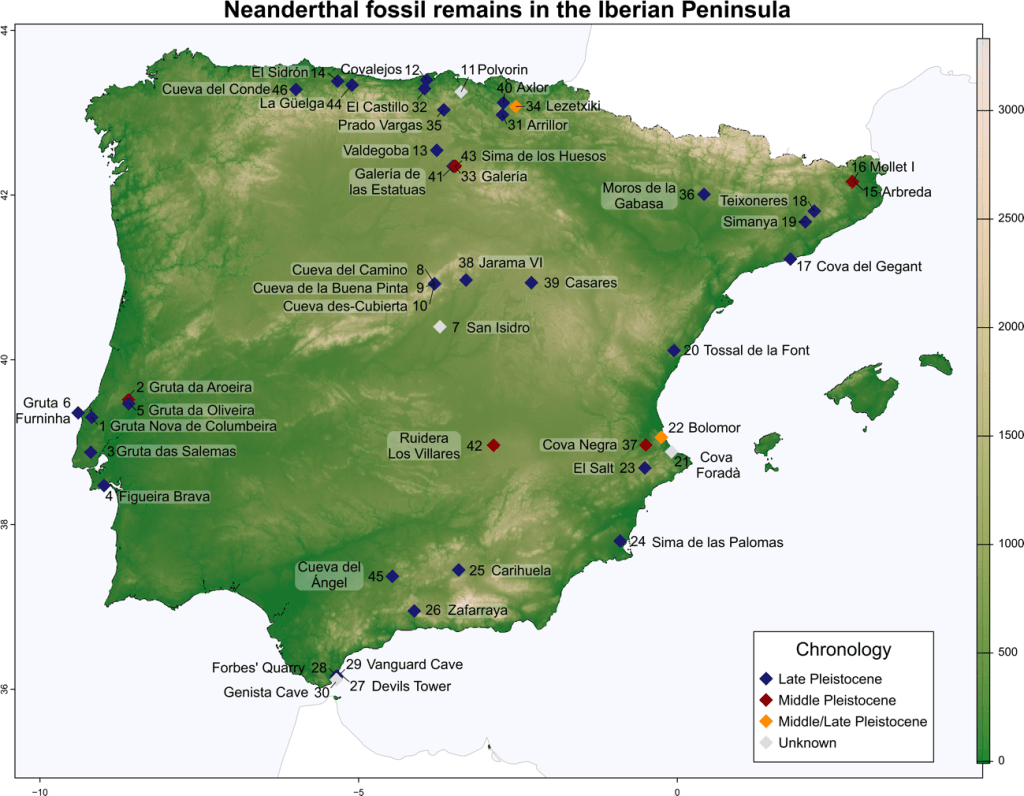
- La revisión del registro fósil de las poblaciones neandertales y preneandertales en la península ibérica indica que, aunque no se han documentado sepulturas formales como en otros lugares europeos, sí existió una notable diversidad espacial y temporal y complejidad en sus prácticas funerarias, como la acumulación intencional de cuerpos en lugares seleccionados en el interior de cuevas (Sala et al., 2025). Esta diversidad no parece estar relacionada con los cambios climáticos ocurridos a lo largo del Pleistoceno.
- Hace 125 ka, neandertales en Neumark-Nord (cerca de Halle, Alemania), transportaron restos de al menos 172 mamíferos (caballos, ciervos y bóvidos) para fracturar los huesos y extraer la grasa de su interior. También se han hallado en el site 16.500 artefactos líticos, restos de elefantes procesados y de fuego en un área de unos 50 m2. Los restos de almidón encontrados sugieren que los neandertales cocinaban en agua, y el calentamiento de alquitrán de abedul sugiere que los huesos fueran almacenados para el procesamiento de la grasa de manera estacional, una práctica de intensificación de recursos que hasta ahora solo se había documentado en el Paleolítico superior (Kindler et al., 2025).
- En la costa noreste del Egeo, Ayvalık es una región en Anatolia occidental que estuvo expuesta intermitentemente como un puente de tierra firme durante los períodos de descenso del nivel del mar en el Pleistoceno (hasta 100 m), lo que generó oportunidades para la ocupación y movilidad humanas tempranas (Bulut et al., 2025). Allí se han identificado 138 artefactos líticos en 10 yacimientos a lo largo de una región de 200 km2. El conjunto más extenso está atribuido al Paleolítico medio, con predominio de Levallois. Hay un pequeño conjunto de herramientas del Paleolítico inferior, como bifaces y cuchillos. El Paleolítico superior y el Epipaleolítico están representados por tecnologías de cuchillos y hojitas.
- La malformación de Chiari tipo I (CM-I) es una protrusión del cerebelo hacia el canal espinal y podría estar vinculada con variantes genéticas heredadas de neandertales. A partir del análisis de morfometría 3D realizado sobre 46 individuos con CM-I y 57 controles, en comparación con ocho cráneos de homininos, Plomp y colaboradores han observado que solo los cráneos con CM-I se asemejaban a los de neandertales. Esta condición produce dolores de cabeza, mareos, problemas de equilibrio y visión borrosa.
- Una nueva investigación de Beasley, Lesnik y Speth sugiere que los neandertales podrían haber comido carne almacenada que ha comenzado a sufrir putrefacción y contiene larvas de mosca y gusanos, como hacen algunas sociedades de cazadores recientes. Este consumo explicaría los niveles altos de isótopos estables de nitrógeno en el colágeno óseo neandertal, lo que les haría parecer hipercarnívoros cuando tal vez no lo eran. Los animales excretan una proporción ligeramente mayor de nitrógeno-14 en la orina que la que ingieren, lo que tiende a concentrar ligeramente el nitrógeno-15 en sus tejidos y tiende a filtrarse a lo largo de la cadena alimentaria. De esta forma, los animales tienen más que las plantas, y los carnívoros más que los herbívoros.
- Se han hallado huellas de neandertales en las playas portuguesas de Telheiro y Monte Clérigo, que sugieren la presencia de bebés en cacerías en grupo hace unos 80 ka. La huella más antigua es una dejada por un adolescente o una mujer joven en una duna de Praia do Telheiro hace unos 82 ka, mientras que en en Monte Clérigo hay otras 10 huellas pertenecientes a tres individuos de hace unos 78 ka (un hombre adulto de entre 1,69 y 1,73 m, un niño de entre 7 y 9 años y un bebé menor de 2). Este hallazgo sugiere que las familias neandertales empleaban estrategias de emboscada en dunas costeras para atrapar presas como el ciervo rojo. En esta organización social compleja, los individuos infantiles también comenzarían a aprender estas habilidades de caza (de Carvalho, Cunha, Belo et al., 2025).
- En Obi-Rakhmat (Uzbekistán), en capas de unos 80 ka, se han identificaron artefactos microlíticos triangulares (micropuntas no retocadas) cuya morfología y fracturas por impacto sugieren que podrían haber sido montados en astas tipo flecha. Esta tecnología, hasta ahora asociada con Homo sapiens del Paleolítico superior, aparece aquí sorprendentemente temprano, aunque los autores no afirman conclusiones firmes (Plisson et al., 2025).
- Hace 70 ka, los neandertales de Axlor (España) ocupaban el lugar estratégicamente y planificaban actividades en función de la disponibilidad de los animales a cazar durante la temporada cálida (Uzunidis, Roussel, González-Urquijo et al., 2025). La cabra montés se cazaba a lo largo de toda la temporada cálida, con picos en primavera-verano y verano-otoño; el ciervo se cazaba sobre todo en verano y principios de otoño; el bisonte en primavera y otoño; y los caballos, rebecos y corzos en verano y otoño, aunque de manera más esporádica. El nivel D de Axlor es particularmente rico en restos arqueológicos, con más de 140.000 fragmentos óseos y unos 11.000 útiles líticos, reflejando una intensa ocupación humana.
- Nuevas dataciones por luminiscencia (OSL) de dos importantes sitios de Olduvai: 153–123 ka para el lecho inferior de Ndutu (extendiendo el rango de las ocupaciones iniciales MSA), 87-71 ka para el lecho superior de Ndutu y 65–33 ka para Naisiusiu (confirmando una llegada temprana de la LSA). La transición entre la MSA y la LSA fue rápida, en torno a 50–45 ka (antes de lo estimado hasta ahora en Olduvai), y permite relacionar estos sitios con Nasera y Mumba (Martín-Perea et al., 2025).
- Allen y O’Connell (2025) defienden que el ADN antiguo hacer cuestionar la cronología de 65 ka sugerida por Clarkson et al. (2017) para la llegada de los humanos modernos a Australia, a partir del registro arqueológico de artefactos líticos y de ocre en Madjedbebe. El argumento básico es que todos los humanos modernos no africanos, incluidos los indígenas australianos, son portadores de ADN neandertal, y los estudios genéticos muestran que neandertales y humanos modernos se cruzaron en Eurasia durante un «pulso» de 7000 años hace entre 50,5 y 43,5 ka, lo que concuerda con la datación de otros yacimientos arqueológicos de 50 ka o menos. Esto no excluye que podría haber más de una llegada temprana de humanos, sin que su ADN haya dejado rastro en el ámbito del estudio realizado (Veth et al., 2025). Además, dado que el arte rupestre de Sulawesi tiene como mínimo 51,2 ka, no sería sorprendente una llegada a Sahul de mayor antigüedad.
- En un trabajo genómico posterior, Gandini y colaborares concluyen que la colonización de Sahul ocurrió hace unos 60 ka, apoyando la “cronología larga” y proponiendo al menos dos rutas de llegada distintas. Llegan a esa conclusión analizando 2456 mitogenomas de poblaciones indígenas de Australia, Nueva Guinea y Oceanía, combinando la reevaluación de la tasa de mutación mitogenómica con datos genómicos, del cromosoma Y, arqueológicos y climatológicos, mostrando que las divergencias genéticas clave se alinean mejor con una llegada temprana (~60 ka). Además, la amplitud y distribución de linajes mitocondriales se explican más coherentemente mediante asentamientos iniciales más antiguos y múltiples rutas de entrada.
- Las cuevas de Amud y Kebara, en el norte de Israel, son yacimientos del Paleolítico medio aproximadamente contemporáneos (70-50 ka), con ocupaciones neandertales que incluyen herramientas líticas, restos de fauna, evidencias de fuego y restos humanos. A partir de similitudes en la fauna hallada, se compararon las técnicas de procesamiento animal mediante el análisis de 344 huesos con marcas de corte. Aunque las especies cazadas son similares, así como su tecnología lítica, se detectaron diferencias notables en la densidad, distribución y forma de las marcas entre ambos sitios. Estos contrastes podrían reflejar diferencias de tradiciones culturales y aprendizaje social entre los grupos neandertales de cada cueva (Jallon et al., 2025).
- Las once campañas paleoantropológicas desarrolladas en Guinea Ecuatorial permiten entender las capacidades tecnológicas y culturales de los humanos modernos en ecosistemas tan hostiles como las selvas tropicales africanas, caracterizados por una densa vegetación, altas temperaturas y recursos impredecibles (Rosas et al., 2025). En 50 de 449 afloramientos estratigráficos investigados se han encontrado evidencias de actividad antropogénica, con 868 herramientas líticas datadas entre 45 y 21 ka, al final del Paleolítico medio africano o MSA (que comprende entre 300 y 25 ka), cuando en otras zonas del continente ya se había desarrollado el Paleolítico superior. Los sitios costeros muestran una mayor diversidad de materia prima y morfotipos similares al Achelense, ausentes en las regiones del interior, donde destaca una amplia variedad de útiles atribuibles al tecnocomplejo Lupemban, el MSA asociado a esta región centroafricana. Este tecnocomplejo parece agregar morfotipos del Modo 2, como bifaces y hendidores, y técnicas y morfotipos de producción de lascas del Modo 3, lo que potencialmente indica un grado de continuidad cultural durante al menos 250 ka.
- Hace 45-70 ka los neandertales de El Castillo (Cantabria, España) usaban para la talla de herramientas materia prima local, pero también adquirida a grandes distancias. Se ha identificado sílex de Tercis (Francia) a 420 km, radiolarita de Alba (100km), sílex de Portillas (133km) y Piedramuelle (246km) en Asturias, de Treviño (169km) en la cuenca del Ebro o Flysch (161km) en el País Vasco (Herrero-Alonso et al., 2025).
- Se ha publicado un taller en Roche-à-Pierrot, Saint-Césaire (Francia) que combina una industria temprana del Paleolítico superior y cuentas de concha, acompañada de pigmentos rojos y amarillos. Esta combinación hasta ahora no estaba documentada. Fechado en al menos 42 ka, se vincula a la cultura Chatelperroniense (Bachellerie et al., 2025).
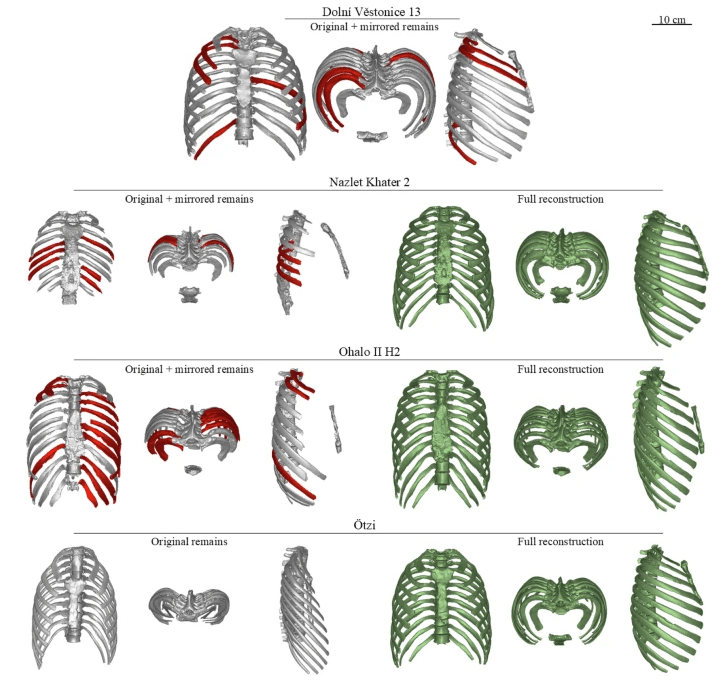
- Investigaciones recientes indican que el tórax esbelto de los humanos modernos es una característica derivada. Para comprobar si los primeros H. sapiens presentaban una morfología torácica primitiva o derivada, se han reconstruido digitalmente las cajas torácicas de Nazlet Khater 2, Ohalo II H2, Dolní Věstonice 13 y Ötzi, y comparado mediante morfometría geométrica con las de 59 humanos modernos y 3 fósiles de otros Homo (Niño de Nariokotome, Kebara 2 y Shanidar 3). Los especímenes fósiles mostraron proporciones torácicas globulares similares a las de los humanos actuales, pero se observó una relación entre morfología torácica y clima: formas más pequeñas y cilíndricas en climas cálidos (Nazlet Khater 2, Ohalo II H2) y más amplias en contextos fríos (Dolní Věstonice 13). Ötzi, por su parte, presentó rasgos mixtos, posiblemente adaptativos a la trashumancia alpina. El estudio revela una mayor complejidad y plasticidad en el tórax sapiens de lo que se había supuesto (López-Rey, Crevecoeur, May et al., 2025).
- Boder y compañeros explican la curiosa predilección por piedras de colores identificada en distintos sitios de Esuatini. Durante la MSA, el jaspe rojo es poco frecuente o está ausente en los conjuntos arqueológicos, mientras que la calcedonia verde y otras variantes de sílex se observan con frecuencia. Esto a pesar de que la fuente de jaspe rojo se encuentra cerca del afloramiento de calcedonia verde. Durante la LSA, el jaspe rojo y una variante de sílex rojo de procedencia desconocida aparecen con mayor frecuencia. Algunas variedades de materia prima fueron transportadas entre 20 y 100 kilómetros de distancia.
- Siguiendo con los colores, Longo y colaboradores han hallado residuos azules (índigo), procedentes de hojas de Isatis tinctoria L., en zonas de desgaste de herramientas de piedra pulida en la cueva de Dzudzuana (Georgia), fechadas en 32-34 ka. Esto abre nuevas posibilidades de uso de herramientas en el Paleolítico superior, tal vez como preparación de tinte o medicina.
- Y en Alemania, también se han identificado rastros de color azul (azurita) en un artefacto de piedra cóncava del yacimiento de Mühlheim-Dietesheim, datados en unos 13 ka (Wisher et al., 2025). Dada la ausencia de azules en el arte paleolítico, los autores sugieren que estos pigmentos podrían haber sido utilizados para la decoración corporal o para teñir telas, actividades que dejan pocos rastros arqueológicos.
- Entre 2013 y 2022, las excavaciones en el sitio gravetiense de Amiens-Renancourt 1 (norte de Francia) dieron como resultado 21 estatuillas femeninas completas o fragmentadas, talladas en creta local (Paris et al., 2025). Destaca el hallazgo en 2021 de una cabeza de 21x13x17 mm, esculpida con el rostro detallado y un peinado o tocado de forma excepcional, ya que los atributos femeninos suelen ser marcados, pero las extremidades y cabezas apenas están representadas. Se aprecian nariz, ojos (con mirada hacia arriba), mejillas y cuello. El tocado o peinado va desde la frente hasta la nuca, con tres tipos distintos de decoración incisa. Su datación es 27,2-27,8 ka. La industria lítica, ósea y objetos de adorno sugieren una intensa actividad en el sitio y organización espacial compleja.

- El cráneo con deformación intencional más antiguo de Europa procede de la cueva Arene Candide, Italia (Mori, Sparacello, Riga et al., 2025), donde se enterraron al menos 22 individuos en el Paleolítico superior. El cráneo AC 12 está datado en 12,6-12,2 ka y presenta un alargamiento del tipo anular, logrado mediante herramientas blandas, como telas apretadas. El otro tipo de alargamiento que suele documentarse desde finales del Pleistoceno es el tabular, logrado mediante objetos rígidos, como tablas de madera, lo que aplana las áreas occipital y frontal y ensancha los parietales.
- Xiang-Chun Ju y colaboradores han estudiado el impacto de la reducción de la actividad de la enzima adenilosuccinato liasa (ADSL), específicamente la mutación A429V, que no está presente en los genomas de neandertales ni denisovanos, pero sí aparece en el 97% de los humanos actuales. En ratones hembra genéticamente alterados, los especímenes con la variante A429V demostraron una mayor capacidad para competir por el agua. Una reducción en la actividad de esta enzima podría haber sido seleccionada por sus ventajas conductuales en condiciones de recursos limitados, aunque no pueden extrapolarse estos hallazgos directamente al comportamiento humano.
- La adaptación de los denisovanos a sus hábitats influyó en la configuración de la resistencia genética humana moderna a las infecciones virales, bacterianas y parasitarias, por ejemplo, afectando a la resistencia a la malaria en poblaciones actuales del sudeste asiático (Trájer, 2025).
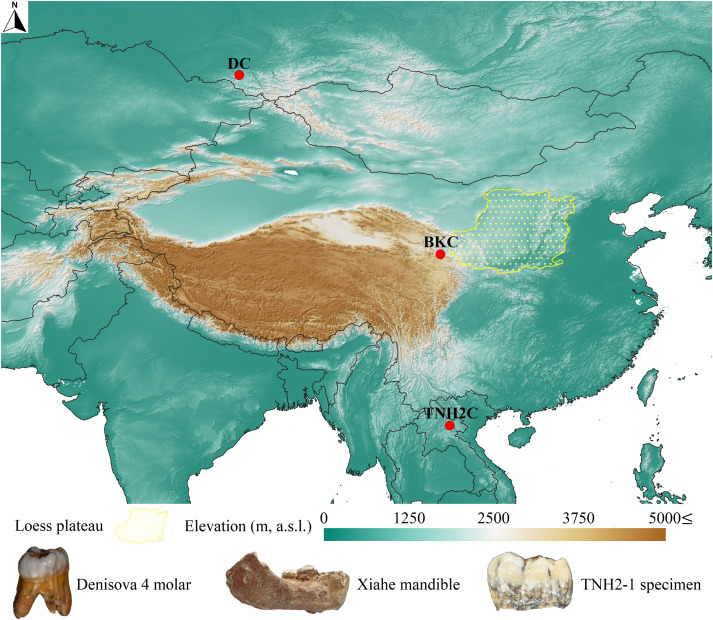
- El gen MUC19 codifica mucinas, glicoproteínas que forman geles con funciones biológicas, como la protección de tejidos. Villanea y colaboradores han encontrado un haplotipo similar al de los denisovanos presente a alta frecuencia en poblaciones amerindias, incluyendo 23 individuos antiguos, todos anteriores a la llegada de europeos y africanos, lo que sugiere una herencia arcaica que fue seleccionada positivamente. Además, han observado que algunos neandertales portaban este haplotipo denisovano, por lo que su transferencia a humanos modernos probablemente ocurrió vía introgresión neandertal y no directamente desde denisovanos.
- Las evidencias más tempranas de momificación de cuerpos secados con humo abarcan un rango de entre hace 12 ka y 4 ka, y proceden de una vasta región que abarca el sudeste asiático y el sur de China. Eran cuerpos momificados sobre todo con posturas agachadas y firmemente atados. Se trata de un conjunto de creencias culturales y prácticas mortuorias que persistieron durante más de 10.000 años entre las comunidades de cazadores-recolectores que estaban relacionadas a través de sus atributos craneofaciales y afinidades genómicas con las poblaciones indígenas de las Tierras Altas de Nueva Guinea y Australia (Hung et al., 2025).
- En la necrópolis megalítica de Los Milanes (Abla, Almería) se han encontrado más de 28.000 fragmentos óseos pertenecientes al menos a 21 individuos que fueron cremados y posteriormente depositados en el interior del monumento. Durante la Edad del Cobre, no solo se empleó la inhumación como ritual funerario, sino también la cremación, que aparece unos 1500 años antes de lo que se pensaba (Becerra Fuello et al., 2025).
- Los desarrollos culturales hacia la agricultura fueron más generalizados de lo que se creía. Hace al menos 9,2 ka, pueblos tan al norte y al este como el sur de Uzbekistán también cosechaban cebada silvestre con hoces (Zhou et al., 2025), muy lejos del Creciente Fértil, donde se sitúa hace unos 10 ka el origen de muchos cultivos clave, como el trigo, la cebada y las legumbres, y la recolección de cereales silvestres por parte de los natufienses.
- Los datos paleogenómicos de 67 individuos prehistóricos ilustra cómo varió la dinámica poblacional entre los cazadores-recolectores indígenas europeos y los agricultores que llegaron a Europa: 1) la mezcla entre ambos grupos aumentó localmente con el tiempo en cada etapa de la expansión neolítica a lo largo de la ruta continental; 2) el tamaño efectivo de la población de agricultores fue aproximadamente cinco veces mayor que el de los cazadores-recolectores; 3) las migraciones esporádicas de larga distancia de los primeros agricultores contribuyeron a su rápida dispersión, mientras que las interacciones competitivas con los cazadores-recolectores fueron limitadas (Tsoupas et al., 2025). Además otro estudio subraya que la presencia de un 10-15% de ascendencia de cazadores-recolectores en los europeos modernos indica una transmisión cultural limitada y un apareamiento predominantemente intragrupal durante la expansión neolítica europea (LaPoste et al., 2025).
- Un trabajo de Hrnčíř, Chira y Gray (2025) plantea que el consumo de alcohol pudo haber favorecido en sociedades humanas complejas la cohesión social, la cooperación, la creatividad y, en el ámbito político, la formación de alianzas y el ejercicio de poder. Con una muestra de 186 sociedades mayoritariamente no industriales, se halló una correlación positiva entre la existencia de bebidas alcohólicas autóctonas y mayores niveles de complejidad política. El alcohol pudo haber contribuido a esta evolución, aunque factores como la agricultura o la religión parecen haber sido motores más determinantes.
- Se ha presentado una nueva evidencia de canibalismo en la Cueva El Mirador de Atapuerca de hace ~5,7 ka (Saladié, Marginedas, Morales et al., 2025). 239 restos humanos tienen marcas de corte, percusión, mordidas y descascarillado. De ellos, 222 tienen signos de haber sido cocinados o expuestos. Los restos son de al menos 11 individuos, y se ha podido estimar la edad de muerte en 7 de ellos: dos <7 años, uno 7-11, uno 14-17, uno 16-17, uno 20-35 y uno >50. El análisis de isótopos indica que era un grupo agrícola de origen local. Este canibalismo no se interpreta como una antropofagia ritual, funeraria o cultural, sino que los humanos fueron descuartizados, cocinados y devorados en un periodo de tiempo breve, posiblemente en el contexto de un enfrentamiento violento entre grupos. Ya se había documentado otro caso de canibalismo en El Mirador, de la Edad del Bronce (4,6-4,1 ka).
