Desconocemos muchos datos de los representantes más antiguos del apasionante género Australopithecus: los Au. anamensis. Pero sí conocemos información de gran valor, y no solo de la morfología que muestran sus fósiles, sino también del contexto ecológico donde vivieron y algunas pistas sobre su modo de vida.
Cómo eran
Al suroeste del Lago Turkana, en el este de África, en 1965 se encontraron fósiles de 4 millones de años en la zona de Kanapoi (Kenia) pero, debido a lo remoto del lugar, no se completó su exploración hasta 1994. La mayoría eran dientes y en total pertenecen a 47 individuos. También se han asignado a este especie un conjunto de restos correspondientes a 31 especímenes en Allia Bay (al este del Lago Turkana) y, recientemente encontrados (2004), 40 dientes aislados en Woranso-Mille (en Afar, Etiopía) y un cráneo casi completo, el primero de esta especie (2016).
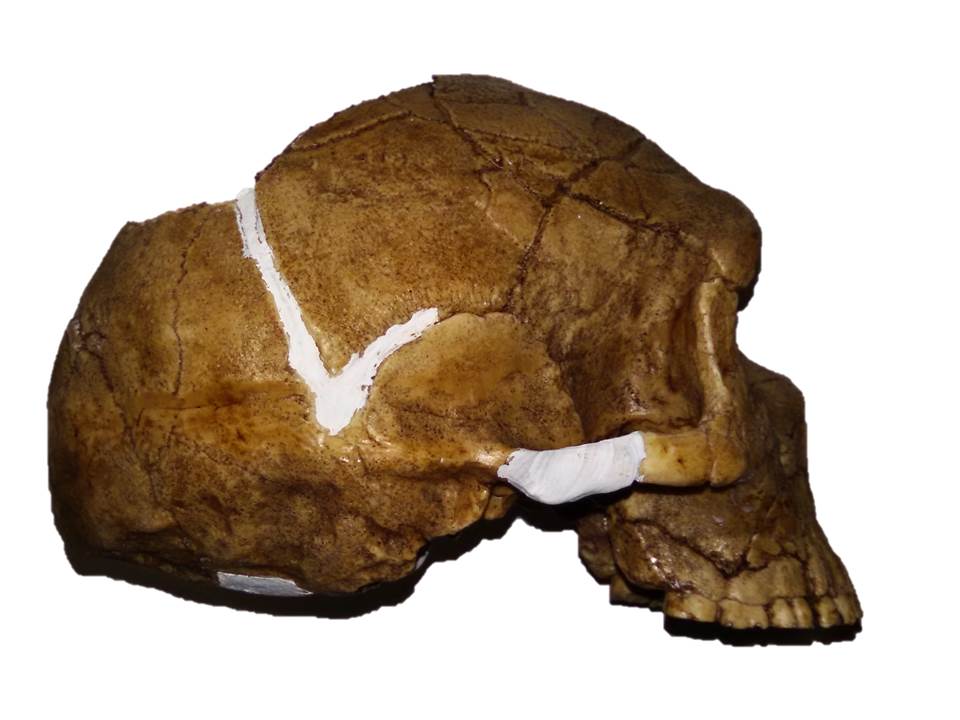
Estos restos constituyen lo que tenemos hasta ahora de la especie Australopithecus anamensis, y están lejos de permitirnos representar un esqueleto completo. En los restos craneales se observan claras reminiscencias primitivas de los simios del Mioceno. El espécimen tipo, KNM KP 29281, es una mandíbula relativamente pequeña con la dentición en forma de “U” estrecha, un ligero diastema entre el canino inferior y el primer premolar (P3) para albergar al canino superior, y una sínfisis mandibular muy inclinada hacia atrás, que presenta en el interior un engrosamiento robusto y alargado. El espécimen KNM KP 29283 es un maxilar con un paladar profundo y caninos con robustas raíces y coronas. Muchos de estos rasgos son primitivos. Del único cráneo (y cara) de esta especie conocido hasta la fecha, MRD, presentado en 2019, escribo en esta otra entrada.

Au. anamensis KNM KP 29283 (I), KNM KP 29281 (C y D). Foto. Roberto Sáez
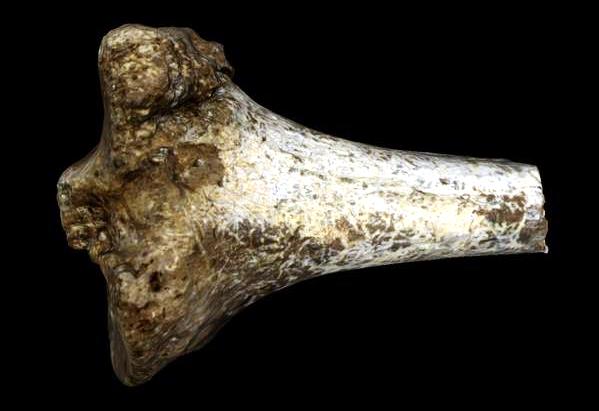
Sin embargo, los restos postcraneales muestran claramente uno de los rasgos clave que nos comenzaron a hacer humanos: la bipedación, manifestada de forma incuestionable por ejemplo en la tibia KNM KP 29285. El extremo que se articula con el fémur es ancho, por tener más tejido esponjoso para absorber las cargas de la locomoción bípeda. El extremo que se articula con el astrágalo tiene un espesamiento que también absorbe el impacto de dicha locomoción. Todos estos rasgos morfológicos los tenemos los humanos.

Au. anamensis KNM KP 29285. Foto: AfricanFossils.org
Su filogenia
Tradicionalmente se pensaba que Au. anamensis probablemente dio lugar a Au. afarensis por anagénesis o evolución filética, que implica la evolución progresiva de una especie hacia otra por cambios en la frecuencia genética de una población entera. Como especie antepasada, también es posible que Ardipithecus ramidus esté relacionada con Au. anamensis.
En 2019 se han revisado algunos materiales asignados con dudas a Au. afarensis pero de antigüedad similar a la de Au. anamensis, y se ha encontrado que probablemente ambas especies convivieron durante unos 100.000 años en la misma zona, lo cual pone en duda la hipótesis de la anagénesis. En todo caso, este estudio también refuerza la posible relación filogenética entre Ar. ramidus, Au. anamensis y Au. afarensis, un interesante y complejo debate que sigue abierto. Doce premolares encontrados en Woranso-Mille en 2014 son un mosaico de caracteres entre Au. anamensis y Au. afarensis, y menos primitivos que A. ramidus.
Su modo de vida
Los yacimientos con presencia de Au. anamensis tienen contextos ecológicos muy similares a los de Au. afarensis, y de hecho representan sistemas muy variados desde bosques abiertos hasta ambientes cerrados de bosques de galería.
Las estrías de la dentición postcanina de algunos especímenes muestran una alimentación propia de sabanas arbustivas similar a la de primates actuales como el babuino y el mono verde. Era abrasiva y se componía de semillas, hojas, cormo y, en mucha menor medida respecto a los Au. afarensis, también frutas. Este estudio apoya el aumento de la robustez de la dentición y del aparato masticador como caracteres evolucionados respecto a los de Ardipithecus ramidus.

Representación de Au. anamensis. Foto: Museum für Naturkunde Berlin
Más información: El nuevo cráneo de Australopithecus anamensis: ¿encaja bien en el «árbol»? | Nutcracker Man