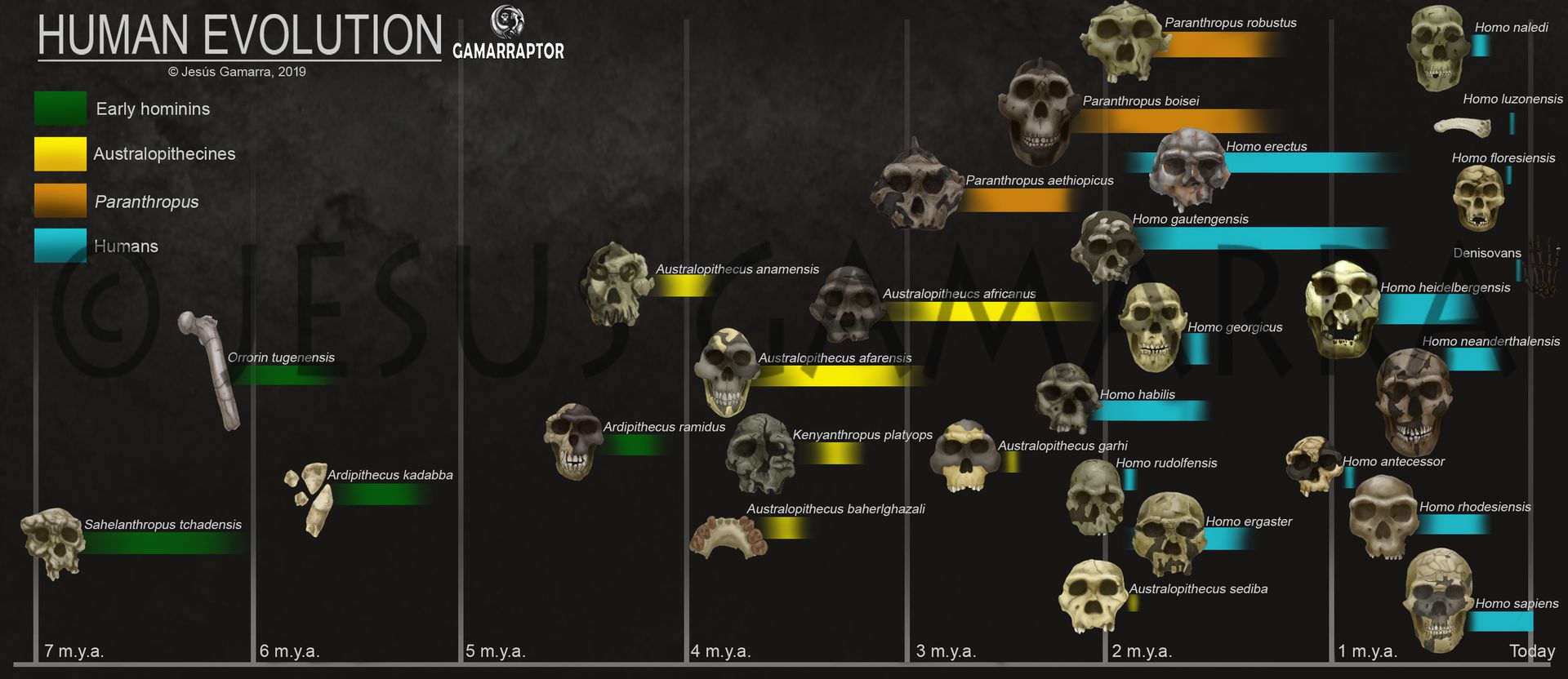
Se ha publicado un nuevo conjunto de dientes aislados encontrados en el yacimiento de Ledi-Geraru, Etiopía (Villmoare, Delezene, Rector et al., 2025). En este lugar también se encontró una mandíbula con dientes asociada a Homo (Villmoare et al., 2015), cuya antigüedad (2,8 Ma, millones de años) le haría ser el fósil más antiguo conocido de nuestro género.
Ledi-Geraru se encuentra al norte de la región de Afar. Esta región es importante para entender la confusa aparición de las primeras formas de Homo en paralelo al desarrollo de Australopithecus entre hace 3 y 2 Ma representados, por ejemplo, por la especie Australopithecus garhi, de 2,5 Ma (Asfaw et al., 1999).
Los rasgos morfológicos de estos nuevos dientes han permitido interpretar algunos como cercanos a los primeros Homo, mientras que otros se aproximan a Australopithecus, sin poder determinarse a qué especie. Estos descubrimientos deben entenderse, ante todo, como datos adicionales que amplían nuestro conocimiento y enriquecen hipótesis previas, como la mencionada existencia muy temprana de primeros Homo en la región contemporáneos a los australopitecos que vivieron durante cientos de miles de años más. Pero estos pocos dientes no suponen una revolución del árbol evolutivo humano ni su reescritura. De hecho, los propios autores de la publicación subrayan la cautela con la que debe leerse su interpretación: ni estos dientes permiten determinar con claridad la aparición del género Homo, ni definir una nueva especie de Australopithecus. Más bien, invitan a seguir explorando la difusa frontera entre Australopithecus y los primeros Homo, aportando piezas nuevas a un puzle todavía muy incompleto, sin caer en las narrativas simplistas sobre «nuevos ancestros humanos» a las que, lamentablemente, estamos habituados.
En este post, he tratado de describir de forma sencilla estos fósiles para ilustrar la interesantísima variabilidad dental que los distingue de otras especies de australopitecos, como Australopithecus afarensis y A. garhi, así como de Paranthropus, mientras que algunos recuerdan más a los dientes humanos. Sigue leyendo