En trabajos anteriores, conocimos las dataciones de las representaciones pictóricas rupestres más antiguas conocidas, realizadas hace más de 45 ka (miles de años) en la isla de Sulawesi (Indonesia). Esas dataciones se obtuvieron mediante el análisis de series de uranio (U-series) de los depósitos de calcita que recubren las pinturas en varias cuevas de piedra caliza de Maros-Pangkep, en el sur de la isla. Más abajo se explican cuáles son esos paneles pictóricos y sus antigüedades.
El método de datación se basa en la medición del decaimiento radiactivo de isótopos de uranio a torio, proceso que ocurre de manera constante a lo largo del tiempo. Dado que el torio no es soluble en agua, no se incorpora a los depósitos minerales desde el entorno, permitiendo que su presencia en la calcita sea exclusivamente el resultado del decaimiento del uranio. Al analizar la cantidad relativa de uranio y torio, se puede determinar la edad del depósito. y, por tanto, la del pigmento que está justo por debajo. Un nuevo trabajo emplea una variación de esa tecnología, series de uranio por ablación láser (LA-U-series), que emplea un láser para quemar capas de calcita de menos de 50 micras de espesor justo encima del pigmento. Con este método se puede estimar con mayor precisión la edad del depósito más antiguo, y por tanto de la pintura. Además, es menos destructivo por requerir muestras más pequeñas.
1) Leang Karampuang: 51.200 años
En este panel de techo existe una gran pintura roja que representa un probable cerdo verrugoso (Sus celebensis) de casi un metro de ancho, en posición estática, con la boca parcialmente abierta. Esta especie de suido es pequeña (de 40 a 85 kg), con patas cortas, verrugas faciales, y muy abundante en Indonesia. Junto a él, hay tres figuras humanas en posición dinámica cerca de la cabeza del cerdo, que tienen aparentemente los brazos extendidos, y dos de ellas sostienen un objeto similar a un palo o una varilla. La datación de la figura del cerdo mediante la nueva técnica mencionada es de al menos de 51,2 ka. Existen dos huellas de manos contemporáneas a la figura. Hay otros cinco cerdos pintados en esta cueva, pero sin relación con el primero.
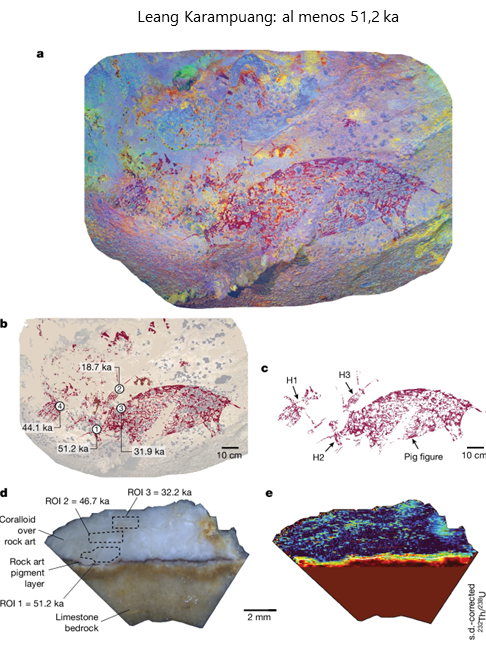
Panel de Leang Karampuang. Crédito: Oktaviana, A.A., Joannes-Boyau, R., Hakim, B. et al. (2024)