
Me había propuesto en este trimestre ser realmente estricto en el número de novedades seleccionadas. Os aseguro que lo he sido, a pesar de lo cual han sido 49 las que recopilo. Estamos en un tiempo fantástico para la paleoantropología.
- Las tasas de cicatrización de heridas humanas son tres veces más lentas que en primates no humanos, las cuales, a su vez, son similares a las de otras especies animales como roedores. Matsumoto-Oda y colaboradores se plantean la desventaja evolutiva que pudo haber supuesto la pérdida del vello corporal. A diferencia de los grandes simios, los humanos experimentaron en su evolución un aumento en la densidad de las glándulas sudoríparas y una disminución en la densidad del vello corporal, acompañado del engrosamiento del tejido subcutáneo como forma alternativa de protección interna. Esto habría contribuido a la evolución de tasas de cicatrización más lentas, que pueden dificultar el acceso a los alimentos, reducir la capacidad de evadir depredadores e influir sobre las necesidades de energía para la reparación de tejidos, el crecimiento y la reproducción. Es posible que el desarrollo del cuidado a individuos mayores y discapacitados entre humanos, así como el uso de plantas medicinales, mitigaran la desventaja evolutiva de la cicatrización tardía de las heridas.
- Los primeros Homo (incluyendo H. habilis y H. rudolfensis) y Paranthropus boisei son propuestos como los fabricantes de útiles olduvayenses (Williams et al., 2025). Los primeros miembros del género Homo siguen siendo los principales candidatos para la fabricación y el uso del Olduvayense a lo largo de su rango temporal más amplio (c. 3,25-1,6 Ma, millones de años) pero, considerando que el Olduvayense pudo darse hasta hace 1,2 Ma, P. boisei sería el principal candidato para la producción de los artefactos posteriores.

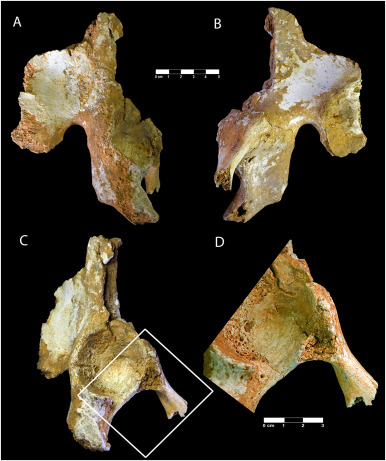
- El espécimen SWT1/HR-2 de Paranthropus robustus, compuesto por un os coxae parcial articulado, el fémur casi completo y la tibia completa, procedente de Swartkrans Miembro 1 (1,7-2,3 Ma), indica bipedación completa y una estatura de solo 1,03 m, el hominino más pequeño conocido (Pickering et al., 2025).
- Tras conocer el sexo biológico del espécimen Sts 63 (Australopithecus africanus) del Miembro 4 de Sterkfontein gracias a su proteoma conservado en un diente (Madupe et al., 2025), ahora han seguido el mismo camino cuatro especímenes de Paranthropus robustus de Swartkrans, de unos 2 Ma: son los dientes SK 830, SK 835, SK 850 y SK 14132. El estudio de la proteína amelogenina ha podido determinar que dos de los dientes tenían la variante específica de los machos (Madupe et al., 2025). Además, el polimorfismo de un solo aminoácido y la variación en la forma de la unión esmalte-dentina indicaron subgrupos potenciales presentes dentro de P. robustus. Anteriormente, se tenía identificada una variante genética única para la proteína polishina en dicha especie, pero este estudio mostró variaciones entre los cuatro individuos analizados. Esto podría reflejar dinámicas poblacionales complejas, microevolución o la presencia de grupos o taxones distintos.
- El estudio de las estructuras dentales y craneales de tres especímenes infantiles de Homo tempranos de 2 Ma revela una gran diversidad de rasgos (Braga y Moggi-Cecchi, 2025). Se trata de la mandíbula Omo 222-1973-2744 atribuida a Homo habilis, y la mandíbula KW 7000 de Kromdraai y el maxilar DNH 83 de Drimolen, atribuidos a Homo aff. erectus. El estudio revela una combinación única de rasgos mandibulares y dentales en estos individuos infantiles, que sugiere una diversidad de trayectorias de desarrollo desde el nacimiento en los Homo tempranos. Los autores sugieren la existencia de ramas evolutivas bien diferenciadas dentro del género Homo, y subrayan la importancia de este factor para la búsqueda del antepasado común de Homo entre los australopitecos.
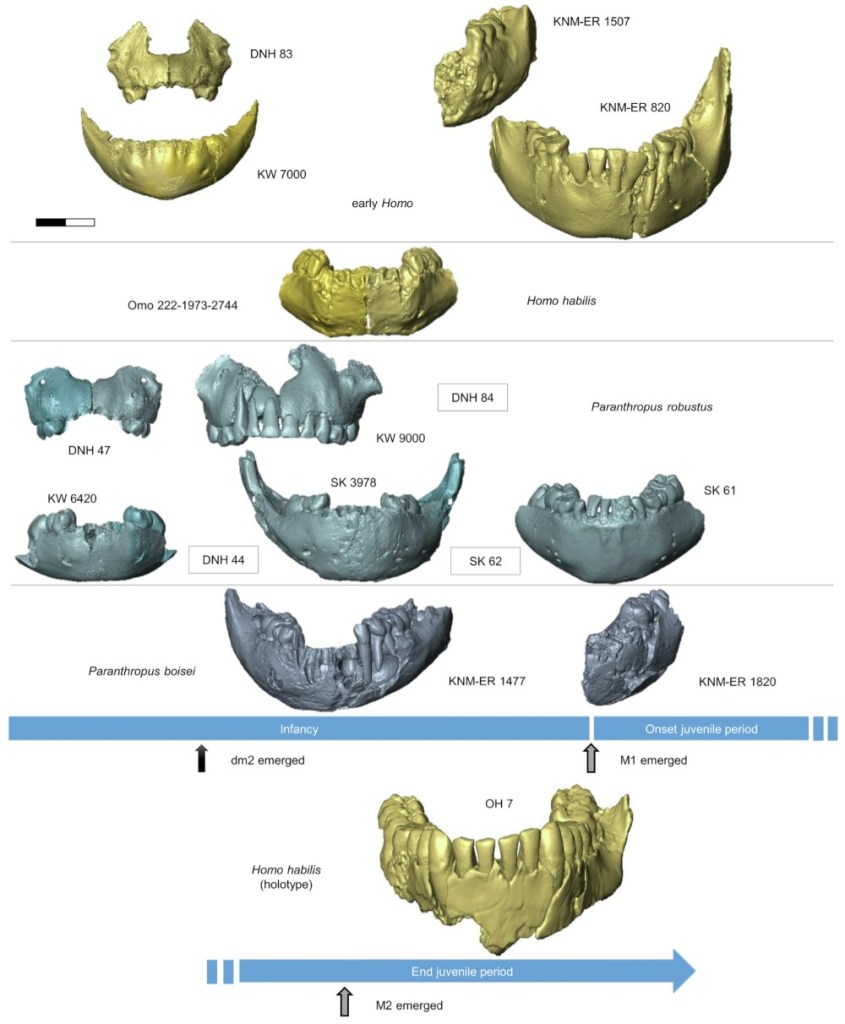
Paranthropus primitivos. Crédito: Braga y Moggi-Cecchi (2025).
- En Paranthropus robustus se conocía una forma particular de cavidades en el esmalte que son uniformes, circulares y superficiales, diferentes de las asociadas con la hipoplasia del esmalte, que son más irregulares y no uniformes. Towle y colaboradores han observado que esta tipología de cavidades se da en los taxones de Paranthropus de África oriental y también en los taxones tempranos de Australopithecus de África oriental. Este hecho puede sugerir una base genética compartida relacionada con la formación del esmalte y factores ambientales o dietéticos, así como ofrecer un fenotipo novedoso para su uso en la clasificación taxonómica de homininos.

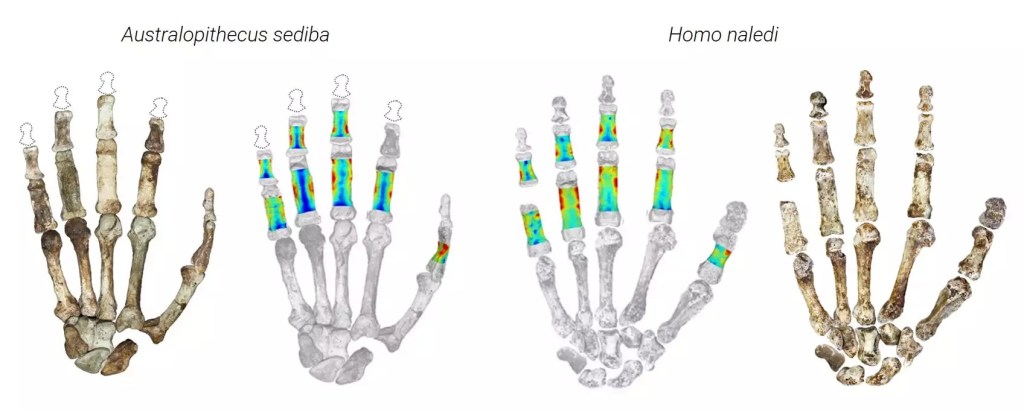
- Un trabajo de Syeda y colaboradores compara las manos casi completas que se conservan de Australopithecus sediba (1,95 Ma) y Homo naledi (250 ka). Las dos especies se adaptaron a sus entornos de distintas maneras, equilibrando el uso de herramientas, el procesamiento de alimentos y la locomoción.
- Mientra tanto, la evolución de la morfología del pie de los primeros homininos parece apoyar la hipótesis de que este rasgo en el Último Antepasado Común de humanos y chimpancés sería tipo Pan (Sekhavati, Prang, Strait, 2025). La estabilidad lateral del pie evolucionó primero en Australopithecus y en homininos posteriores, siendo las primeras sinapomorfías del pie relacionadas con la eversión del pie y del tobillo y la estabilidad medial. El antepasado común de Paranthropus y de Au. africanus + Au. sediba tendría adaptaciones a la vida arbórea.
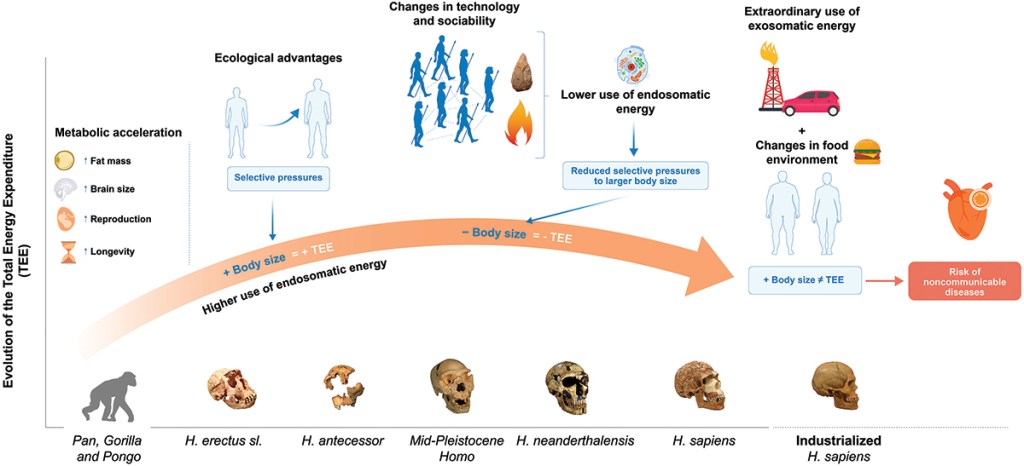
- Hasta ahora, las estimaciones publicadas del gasto energético de los homininos asociado a sus niveles de actividad física habían supuesto valores para los cazadores-recolectores actuales y otras poblaciones con economías de subsistencia de 2 o 2,5 veces el gasto de un atleta de resistencia actual. Un estudio de Prado-Nóvoa y colaboradores afirma que el gasto energético diario estaría sobreestimado en unos 8,4 MJ/día. Los autores han reevaluado el gasto energético en el género Homo de acuerdo a ecuaciones predictivas no basadas en los niveles de actividad, sino en la masa corporal de 112 individuos (H. erectus sensu lato, H. antecessor, otros Homo del Pleistoceno medio, H. neanderthalensis y H. sapiens). El estudio sostiene que durante los últimos 2 Ma probablemente se haya producido una aceleración y desaceleración metabólica vinculada a los cambios en la masa corporal. Estos cambios podrían estar relacionados con la capacidad de ciertas especies para utilizar diferentes fuentes de energía internas y externas (fuego, herramientas). Con ello, se concluye que no debemos asumir que nuestros antepasados vivían en su límite metabólico todos los días (como «máquinas incansables»), sino que la evolución trata de minimizar los costos energéticos innecesarios («aprende a ahorrar»). Nuestra historia metabólica es compleja, adaptativa y eficiente.
- Veníamos conociendo fases húmedas en la llamada «Arabia Verde» durante los últimos 1,1 Ma. Pero hubo más periodos húmedos recurrentes en el interior de Arabia central durante los últimos 8 Ma, de acuerdo al registro climático de espeleotemas desérticos (Markowska, Vonhof, Groucutt et al., 2025). Este es un aporte muy interesante para incorporar en futuros estudios sobre las migraciones de los homininos.
- La conservación de la carne y la grasa y la protección frente a depredadores serían los primeros motores para el uso del fuego por parte de Homo erectus durante el Paleolítico inferior (1,9-0,78 Ma), y no tanto no la cocción de alimentos (Ben-Dor y Barkai 2025). La proyección de datos etnográficos al Paleolítico inferior muestra que la caza de presas de más de 100 kg generaría un rendimiento energético 10 veces mayor que la recolección de plantas (16269 ca/h frente a 1443 ca/h). Presas como un hipopótamo pueden dar alimento a un grupo de 25 individuos durante hasta 22 días. Los primeros yacimientos con restos de fuego cuentan con abundantes restos de fauna, que indica un consumo prolongado en el tiempo. En cambio, la cocción ofrecería ganancias energéticas modestas (~1200 ca/h) insuficientes para compensar el trabajo del mantenimiento del fuego.
- Se ha documentado en el sitio Medzhibozh A (Ucrania) el uso más antiguo conocido del marfil como material para la fabricación de herramientas, en total 24 fragmentos de hace unos 400 ka. De los 24 fragmentos, 11 tienen signos de modificaciones intencionales, 6 de talla, 3 fueron usados para talla bipolar sobre yunque, 2 lascas y 1 punta y 1 núcleo (Stepanchuk y Naumenko, 2025).

- La comparativa del espesor del esmalte en los premolares permanentes de H. antecessor (0,9 Ma) y de los preneandertales de Sima de los Huesos (SH) (0,4 Ma) sugiere: 1) una gran variabilidad intrapoblacional en este rasgo entre los homininos de Atapuerca; 2) la aparición temprana de esmalte reducido en poblaciones europeas; 3) la reducción general de la corona podría haber contribuido a la variación en el grosor del esmalte en estas poblaciones. En SH, los premolares superiores tienen esmalte fino como en neandertales, mientras que en los inferiores es más grueso y similar a H. sapiens. En H. antecessor, el grosor del esmalte en los premolares superiores es muy variable, pero en los inferiores tiende a ser fino como en los neandertales. El grosor del esmalte por sí mismo no es una condición primitiva o derivada (Martín-Francés et al., 2025).
- Durante unas operaciones de dragado en el estrecho de Madura, costa de Java, Indonesia, se han encontrado restos fosilizados de 36 especies de vertebrados, incluyendo dos fragmentos de cráneo de H. erectus (Bello y Stringer, 2025). Su datación por OSL es de entre 162 y 119 ka. Por entonces, el nivel del mar era unos 100 metros más bajo que el actual. La fauna asociada indica que en esa zona H. erectus cazaba activamente bóvidos sanos y fuertes, una práctica que no se había observado en poblaciones anteriores de esta especie en Java. Recordemos que en el valle de Solo ya se habían encontrado 11 cráneos parciales, incluyendo el espécimen más reciente de Java, de hace unos 112 ka. El famoso cráneo del «Hombre de Java» hallado por Eugène Dubois también proviene de la zona, pero antes de la formación del río Solo.
- A partir de 103 rasgos culturales surgidos en el Paleolítico, Coàge y d’Errico evalúan la evolución de la transmisión cultural durante los últimos 3,3 Ma, junto con 24 expertos. Entre dichos rasgos se incluyen tipos específicos de herramientas de piedra, ornamentos como cuentas, pigmentos y prácticas mortuorias como enterramientos y la construcción de túmulos. Los autores identificaron cuándo apareció cada rasgo por primera vez regularmente en el registro arqueológico, lo que puede reflejar que era una práctica común. En cuanto al aprendizaje, analizaron tres aspectos: 1) Espacial: ¿se puede aprender la habilidad observando a distancia o se necesita estar lo suficientemente cerca como para tocar? 2) Temporal: ¿es suficiente una lección corta o se necesitan varias sesiones, quizás centradas en diferentes pasos? 3) Social: ¿quién aprende de quién? Como resultados, plantean que hace 600 ka ya existirían métodos eficaces de transmisión de conocimiento explícita, a través de gestos y expresiones verbales, coincidiendo con la aparición de técnicas como el enmangado, mientras que el periodo 200-100 ka supuso un punto de inflexión crucial para el surgimiento del lenguaje moderno, coincidiendo con la aparición de conductas como el enterramiento.
- Conocíamos la evidencia del lago Schöningen como zona de caza de emboscada, donde homininos cazaron recurrentemente grupos de caballos y allí dejaron múltiples lanzas y otros útiles de caza que se han conservado excepcionalmente. La antigüedad de las icónicas lanzas de madera de Schöningen venía estimándose en unos 300 ka. La datación por termoluminiscencia (TL) del sílex calentado en el sitio 13I-1 era de unos 321 ka, mientras que la datación por luminiscencia ópticamente estimulada (OSL) del sitio 13II-2c1 daba unos 300 ka. Sin embargo, las muestras datadas de las unidades 13I-1 y 13II-2c1 están ubicadas estratigráficamente por debajo de la unidad 13II-4, de donde provienen los objetos de madera y, por tanto, solo sirven como estimaciones de edad máxima. Una nueva datación (Hutson et al., 2025) por geocronología de aminoácidos de múltiples tipos de fósiles recuperados de entre las concentraciones más densas de 13II-4, dan una edad de unos 200 ka para las famosas lanzas de Schöningen. Este método de datación se basa en la degradación dependiente del tiempo de las proteínas dentro de los fósiles.
- La reevaluación del cráneo Maba 1 muestra una combinación de rasgos que se observan en diversas especies, lo que hace imposible una propuesta de clasificación en un taxón hominino. Aunque su cara recuerda a la neandertal, la morfología de su endocráneo está más cercana a la de H. erectus. Maba 1 es un buen ejemplo de la gran variabilidad morfológica entre los homininos asiáticos del Pleistoceno medio (Hui, Wu, Balzeau, 2025).
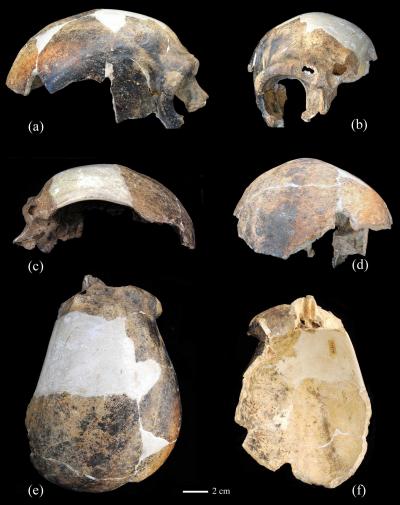
- Un preprint de Feng y otros (2025) propone el cráneo de Yunxian 2 como miembro basal del clado formado por Homo longi y los denisovanos, y se sitúa próximo al último antepasado común de dicho clado y del clado de H. sapiens.
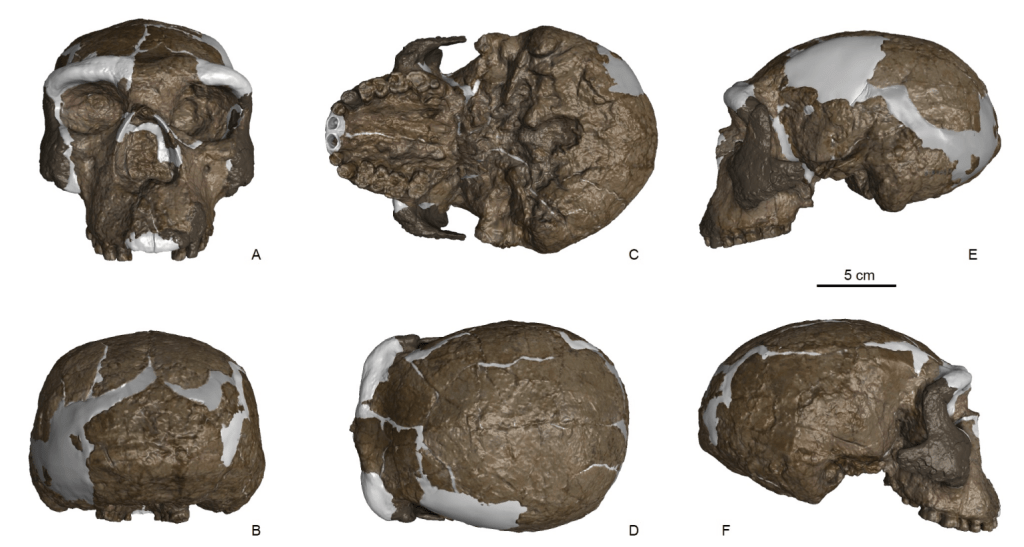
- Poco a poco vamos completando la historia de lo que ocurrió en la cueva Denisova desde hace 300 ka (miles de años). Jacobs, Zavala, Li y otros compañeros describen de forma exhaustiva en su publicación la cronología de los homininos y la fauna que ocuparon la cueva en distintos momentos hasta la llegada de los humanos modernos.
- Mientras tanto, se ha logrado identificar el cráneo de Harbin como denisovano. Por una parte, el ADN mitocondrial obtenido a partir del cálculo dental se sitúa dentro de la variación genética de los denisovanos. Además, el estudio paleoproteómico a partir del hueso petroso también respalda su identidad denisovana.
- Un trabajo de Coutinho-Nogueira y colaboradores analiza si el icónico cráneo Qafzeh 9, de unos 92 ka, es un buen representante de los fósiles del yacimiento de Qafzeh y de los primeros humanos modernos. El nuevo estudio concluye que su morfología craneal difiere notablemente de la de otros individuos adultos del yacimiento, especialmente de Qafzeh 6 y 25. Anteriormente, también se había observado una variabilidad significativa entre los fósiles de Qafzeh en la morfología del oído interno. También muestra variantes anatómicas, rasgos dentales y patologías relacionadas con el crecimiento que no se observan en la mayoría de los demás fósiles de Qafzeh (excepto Q10, enterrado junto con Q9). Tampoco ayuda en las comparativas que se trate de un individuo relativamente joven y que su sexo siga siendo objeto de debate. Por tanto, aunque se trata del fósil mejor conservado del yacimiento, debe cuestionarse la consideración de un representante único del mismo.
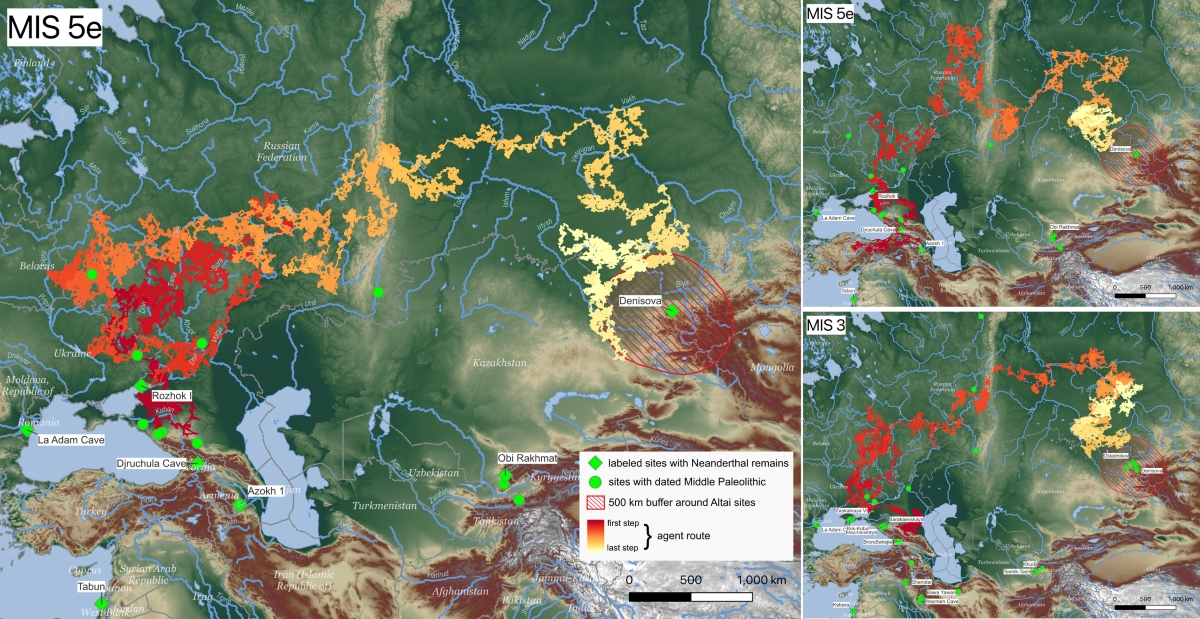
- Los datos genéticos indican que los primeros neandertales probablemente llegaron a Asia poco antes del MIS 6 (190-130 ka), con una fase de dispersión e intercambio genético entre neandertales europeos y siberianos durante MIS 5 (120-80 ka), por ejemplo, observada por las similitudes genéticas entre los neandertales de Mezmaiskaya y Chagyrskaya, y un segundo gran desplazamiento de neandertales desde Eurasia occidental hacia Eurasia central y oriental en algún momento del Pleistoceno tardío. El registro arqueológico proporciona una fecha de llegada al Altái en el MIS 3 (60 ka). Ahora, una simulación ha determinado las probables rutas de migración en los momentos más cálidos y adecuados. Coco e Iovita concluyen que los neandertales probablemente siguieron una ruta septentrional a través de los Urales y el sur de Siberia en todos los escenarios climáticos, y los que partieron del sur o del norte del Cáucaso llegaron al Altái, recorriendo 3250 km en unos 2000 años, durante dos ventanas temporales de clima templado: en el MIS 5e (el Último Interglaciar hace unos 125 ka) y en el MIS 3. Este último coincide con la presencia datada de neandertales en las cuevas de Chagyrskaya y Okladnikov en el Altái.
- La cueva de Valdegoba (Burgos, España) presenta un conjunto de restos de aves consumidas tanto por depredadores rapaces como por humanos (Alonso Caño et al., 2025). Estos fueron grupos neandertales que ocuparon la cueva entre el MIS 3 y el 5c. Se trata de una evidencia más de las actividades de caza y procesamiento de presas pequeñas, como aves, observadas en el comportamiento neandertal, también para adornos personales en otros yacimientos.
- Ha sorprendido un guijarro procedente del abrigo rocoso de San Lázaro (Segovia, España) que presenta modificaciones antropogénicas y una huella neandertal hecha con ocre.

- La aplicación de modelos de distribución de especies para cuantificar los cambios en el nicho bioclimático durante los últimos 120 ka sugiere que, antes de la «gran expansión» fuera de África hace 50 ka, los humanos modernos habían explorado diversos tipos de hábitat, desde bosques hasta desiertos áridos. Los que se dispersaron tenían una flexibilidad ecológica y adaptabilidad distintivas (Hallett, Leonardi, Cerasoni et al., 2025).
- Se ha identificado industria tecnológica Quina en Longtan, suroeste de China, datada en unos 55 ka (Ruan et al., 2025). Se trata de un conjunto completo que exhiben todo el concepto de Quina: explotación de núcleos, producción de lascas grandes y gruesas, y modelado y mantenimiento de raspadores. Este conjunto es contemporáneo a las tecnologías análogas del Paleolítico medio europeo desarrolladas por grupos neandertales adaptados a las oscilaciones climáticas durante el MIS 4 y principios de MIS 3, lo que desafía la visión de que no hay una industria del Paleolítico medio en esta región, revela una diversidad tecnológica en este periodo, y abre la incógnita de si están representando soluciones culturales convergentes, o bien un grupo de neandertales errantes en China.
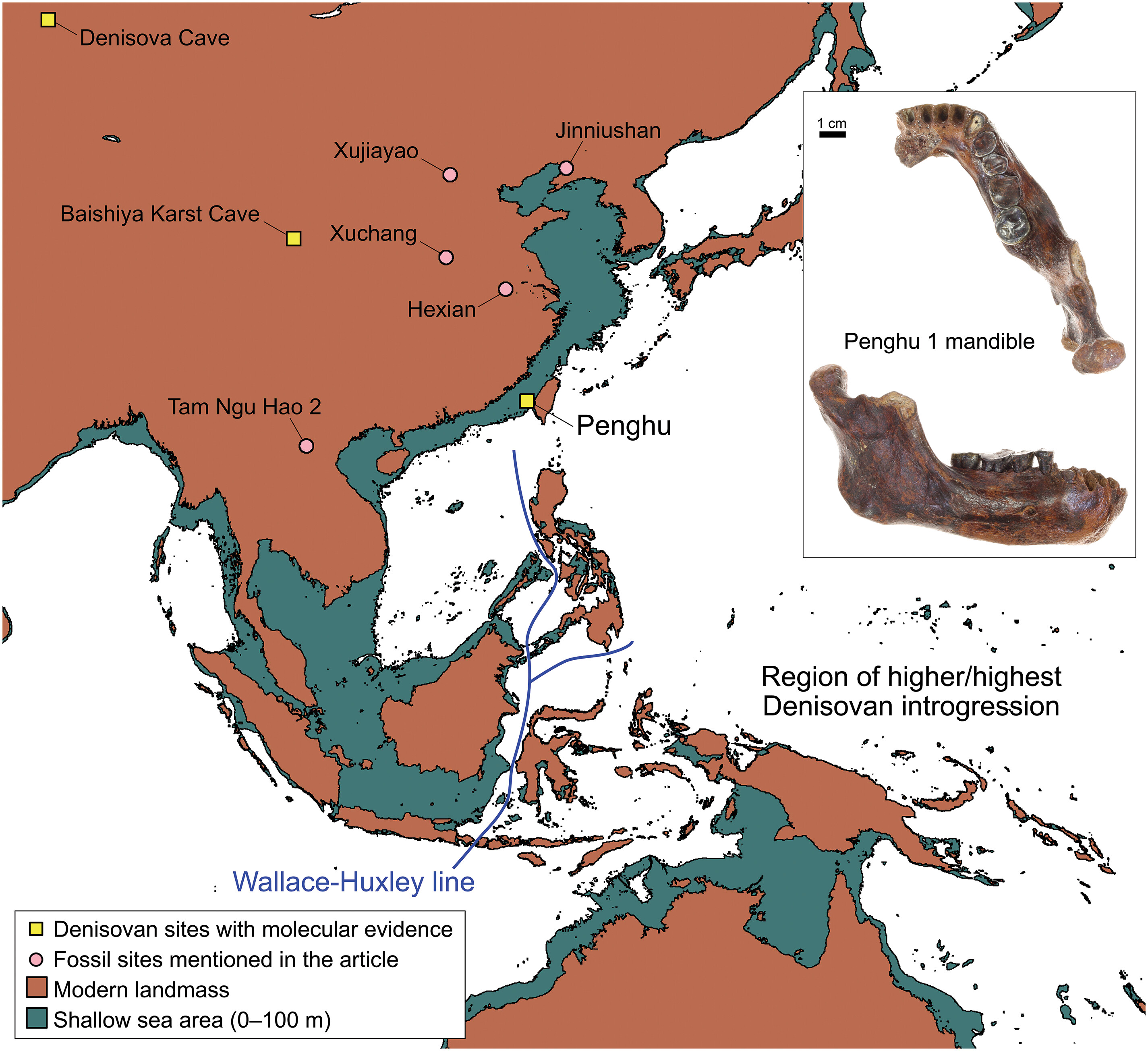
- La mandíbula de Penghu fue recogida en 2008 por pescadores en un canal submarino homónimo, a unas 15 millas de la costa. El estudio de 4241 restos de aminoácidos de 51 proteínas recuperadas de Penghu 1 ha permitido identificar dos variantes específicas de los denisovanos (Tsutaya et al., 2025). El análisis filogenético también agrupa la mandíbula con los denisovanos. Este hallazgo muestra cómo los desinovanos se desplazaron hacia el sudeste asiático, amplía su área de presencia (entre la Cueva Denisova a Penghu hay más de 4000 km) y ayuda a explicar el rastro de su genoma en el de los melanesios actuales.
- Los humanos auriñacienses (42-34 ka) y gravetienses (34-24 ka) usaban adornos similares tanto a escala europea como regional, según describen d’Errico y otros a partir del estudio de enterramientos de ambas culturas. Los ornamentos personales completamente tallados presentan mayores diferencias entre tecnocomplejos, que los producidos a partir de formas naturales mínimamente modificadas. Además, la variabilidad de los ornamentos auriñacienses sugiere conjuntos culturales más fragmentados en comparación con el Gravetiense, lo que implica redes simbólicas más extensas en este último.
- Esta sorprendente herramienta de 72 cm de largo tipo bumerán, hecha con marfil de mamut, procede del nivel VIII de la cueva de Obłazowa (Polonia) y su antigüedad estimada es 39-42 ka, que es la datación de 14 huesos de animales y humanos encontrados en asociación con el artefacto. Como no se encontraron otras herramientas cerca, los autores asumen que el bumerán fue llevado por cazadores-recolectores migratorios. Se emplearía para cazar, cavar, raspar y, dado que tiene restos de pigmento rojo, también pudo haber tenido una función simbólica o ritual (Talamo, Casaccia, Richards, Wacker, Tassoni, Nadachowski, et al., 2025)

- La datación directa de restos de lodo antiguo asociados a las huellas de White Sands confirman su asombrosa antigüedad de entre 20,7-22,4 ka (Holliday et al., 2025).
- Los humanos fabricaban útiles con huesos de ballena hace 20-14 ka, según revela un análisis de 173 herramientas óseas procedentes de distintos yacimientos del Magdaleniense alrededor del Golfo de Vizcaya (McGrath, van der Sluis, Lefebvre et al., 2025). Se han identificado al menos cinco especies distintas de ballena, incluyendo cachalotes, rorcuales y ballenas azules, todas ellas aún presentes en el Golfo de Vizcaya, así como ballenas grises, que en la actualidad viven en el Pacífico Norte y en el Ártico. La máxima difusión y diversidad de productos de ballena se dio hace 17,5–16 ka. Además, se han identificado dos patrones de uso diferentes: 1) como materia prima de grandes dimensiones que permite la fabricación de elementos de caza; 2) transporte de los huesos sin trabajar a los lugares de habitación, tal vez como combustible óseo. Finalmente, aunque la muestra arqueológica incluye solo huesos y objetos óseos, su presencia muestra que los cazadores-recolectores también tenían acceso a otros recursos balleneros, como la piel, la grasa y la carne de ballena (para usos dietéticos y técnicos) y a las placas barbadas (material fuerte y flexible con muchos usos prácticos).
- Se ha documentado un objeto de 9 cm interpretado como probable punta de lanza fabricada en hueso (parece que de bisonte), que habría estado montada en un mango hecho de madera con la ayuda de brea, hallada en un nivel del Paleolítico medio de la cueva Mezmaiskaya en el Cáucaso Norte de Rusia. Su antigüedad estimada en torno a 70-80 ka le hace ser la más antigua conocida en Europa (Golovanova, Doronichev et al., 2025) superando a la de Abric Romaní, que tiene 50 ka (Mateo-Lomba et al., 2024). El contexto indica su asociación con los neandertales de la tradición cultural llamada Micoquiense oriental. Muestra un nivel incipiente de desarrollo de esta tecnología en comparación con las introducidas en Eurasia por los humanos modernos.

- Las poblaciones de humanos modernos en la región china de Yunnan (sudeste asiático) se diversificaron hace menos de 40 ka y presentan ascendencias basales asiáticas que fueron reemplazadas en gran medida por un único linaje, que se diversificó en las ascendencias presentes en la mayoría de los asiáticos actuales. Además, un individuo de Yunnan de 7,1 ka muestra está relacionado con una ascendencia fantasma profundamente divergente que contribuyó a las poblaciones de la meseta tibetana (Wang et al., 2025).
- Los cuerpos de dos mujeres adultas de hace 7 ka procedentes del abrigo rocoso de Takarkori, sur de Libia quedaron momificados de forma natural gracias a la aridez y las altas temperaturas. El estudio genético de sus restos y su comparación con el genoma de unos 800 humanos actuales y 117 africanos de diferentes épocas pasadas, indican que esas mujeres pertenecían a una población sin parentesco con el resto del África subsahariana, que permaneció aislada del sur y del norte (Salem, N., van de Loosdrecht, M.S., Sümer, A.P. et al., 2025). El origen de su linaje se remonta a hace unos 50 ka, cuando su grupo se separó de los humanos modernos que salieron de África en esa época, antepasados de los humanos euroasiáticos actuales. Tenían vínculos con la población de Taforalt (Marruecos) de hace unos 15 ka, mostrando una continuidad genética en el norte de África. La gente de Taforalt tenía un 60% de ascendencia natufiense y un 40% de Takarkori. Los autores sugieren también que el pastoralismo se expandió por el denominado «Sáhara Verde» hacia el sur probablemente por intercambio cultural y no por grandes migraciones, dada la escasa presencia de ADN levantino y la baja proporción de ADN neandertal (0,15%) en los individuos de Takarkori. Esta población habría estado aislada durante milenios tal vez por la segmentación ecológica del Sáhara en hábitats fragmentados durante el periodo húmedo africano, desde hace 14 ka, cuando las lluvias monzónicas crearon el Sáhara Verde tras la última glaciación. Takarkori era una sabana cubierta de pastos, árboles, lagos y ríos habitados por jirafas, hipopótamos y otra megafauna. Este paisaje está representado en las pinturas rupestres de las montañas Tadrart Acacus de Libia, de unos 12 ka.

- La pelvis neandertal El Sidrón SD-1663 presenta rasgos similares con las de otros neandertales, pero también afinidades con las de Homo anteriores, como con el espécimen ER 3228 de H. erectus. Esto sugiere un rango más amplio de variación pélvica dentro de los neandertales de lo que se conocía previamente (Torres-Tamayo et al., 2025).
- Se han estudiado nuevas implicaciones para la interpretación del método Levallois, concretamente las variaciones en el ángulo de impacto del martillo, que tendría un efecto mayor en la formación de lasca de lo que se pensaba anteriormente. Estos resultados sugieren un mayor grado de control cognitivo de lo que se creía previamente. Los talladores neandertales conocerían los ángulos de impacto para controlar el tamaño, la forma y la terminación de las lascas. Desprender lascas más grandes en ángulos bajos requería mayor fuerza y conllevaba un mayor riesgo. Por tanto, la producción de Levallois implicaba más que la preparación del núcleo, requiriendo planificación, coordinación física, toma de decisiones de riesgo y control receptivo de la fuerza (Lin et al., 2025).
- Siete piezas de ocre fueron modificadas hace 70-90 ka en Blombos (Sudáfrica) deliberadamente para retocar artefactos líticos de MSA, mostrando la multifuncionalidad del ocre y la posible importancia personal, cultural o tecnológica de estas herramientas (Velliky et al., 2025).
- El registro de carbono pirogénico depositado en sedimentos del Mar de China Oriental durante los últimos 300 ka sugiere un rápido aumento de la actividad de incendios desde hace aproximadamente 50 ka. Jiang y colaboradores proponen que la intensificación de la expansión global de los humanos modernos, junto con la creciente demanda de uso del fuego durante los períodos fríos, resultó en un aumento significativo en la utilización del fuego a partir de ese umbral de 50 ka.
- Hace 41 ka ocurrió el llamado evento de Laschamps, en que los polos magnéticos de la Tierra se invirtieron y desplazaron hacia el ecuador y el campo magnético había quedado reducido a un 10% del nivel actual. Mukhopadhyay y colaboradores concluyen que H. sapiens se adaptaría y protegería de la radiación solar extrema en Europa y el norte de Asia gracias al uso de ropa adaptada y ocre como protector solar y un incremento del uso de cuevas, mientras que este evento habría contribuido a la extinción de los neandertales. Un evento de este tipo perturbaría la tecnología contemporánea, incluyendo las comunicaciones y la infraestructura satelital.
- Se ha anunciado en prensa el hallazgo de huellas humanas dejadas en nueve lugares de la Cueva del Tesoro de Rincón (Málaga, España). En espera de los resultados de las muestras enviadas para datar, su antigüedad por ahora se estima en unos 40 ka.

- Hace 30 ka, la migración marítima a las Islas Ryukyu de Japón fue una de las travesías marítimas más difíciles logradas en el Pleistoceno superior. Un trabajo de Chang y colaboradores combina simulaciones basadas en tres modelos oceánicos de las condiciones necesarias para cruzar entre Taiwán y la isla Yonaguni, donde actúa la fuerte corriente oceánica Kuroshio, con un viaje experimental. Los autores concluyen que aquel viaje habría sido posible con una canoa escogiendo el lugar de partida, conociendo cómo navegar en la Kuroshio y técnicas de remado y otras habilidades expertas de navegación.
- Nuevas dataciones con series de uranio-torio del Techo de los Policromos de la cueva de Altamira (España) confirman el origen auriñaciense para un claviforme (>32,79 ka) y arrojan una antigüedad mínima sorprendente de 22,6 y 32 ka para las muestras de pigmento tomadas en dos figuras de caballos, lo que les situaría en el Gravetiense o incluso también en el Auriñaciense, apuntando a una coexistencia de los estilos figurativo y simbólico (Shao et al., 2025)

- Se ha publicado una de las evidencias más antiguas que se conservan de marcas por impacto de flecha o proyectil (Sparacello, Thun Hohenstein, Boschin et al., 2025). Procede de Riparo Tagliente (Italia). El fémur y la tibia del individuo Tagliente 1 muestran una clara evidencia de impacto por proyectil desde diferentes direcciones. Su datación directa es de 17-15,5 ka.
- En Montlleó, sitio pirenaico a 1100 m de altura, se ha documentado una larga secuencia de ocupación humana que comienza en el Último Máximo Glacial y termina después del mismo, abarcando entre hace 23 y 17 ka, correspondiente a tres tradiciones culturales distintas (Sánchez de la Torre et al., 2025).
- El estudio genómico de 2762 individuos ilustra 50.000 años de historia evolutiva de la India (Kerdoncuff et al., 2025). 1) La mayoría de los indios poseen ascendencia principalmente de tres grupos ancestrales: cazadores-recolectores del sur de Asia, pastores esteparios euroasiáticos y agricultores neolíticos relacionados con las culturas iraníes y de Asia central. 2) El alto nivel de parentesco indica fuertes eventos fundadores con tendencia hacia la endogamia. 3) La mayor parte de la variación genética en los indios proviene de una única gran migración desde África que ocurrió hace unos 50 ka, con flujo genético del 1 % al 2 % proveniente de neandertales y denisovanos. Los indios exhiben la mayor variación y poseen la mayor cantidad de segmentos de ascendencia neandertal específicos entre los humanos actuales. 4) Las variantes introgresadas de los neandertales y los denisovanos contribuyen a la respuesta inmunológica innata.
- Otro estudio genómico, esta vez abarcando a 1537 individuos de 139 grupos étnicos (Gusareva et al., 2025) revela una historia de migración humana del norte asiático hasta Sudamérica: la ascendencia siberiana occidental, representada por los kets y los nenets, contribuyó a la ascendencia genética de la mayoría de las poblaciones siberianas. Los beringianos occidentales, incluyendo a los koryaks, inuit y luoravetlans, muestran una adaptación genética al clima ártico. En Sudamérica, los primeros migrantes se dividieron hace unos 13,9 ka en cuatro grupos: amazónicos, andinos, amerindios del Chaco y patagónicos. Su migración más larga condujo a una disminución poblacional, mientras que el asentamiento en los diversos entornos de Sudamérica causó un aislamiento espacial que redujo la diversidad genética e inmunogénica.
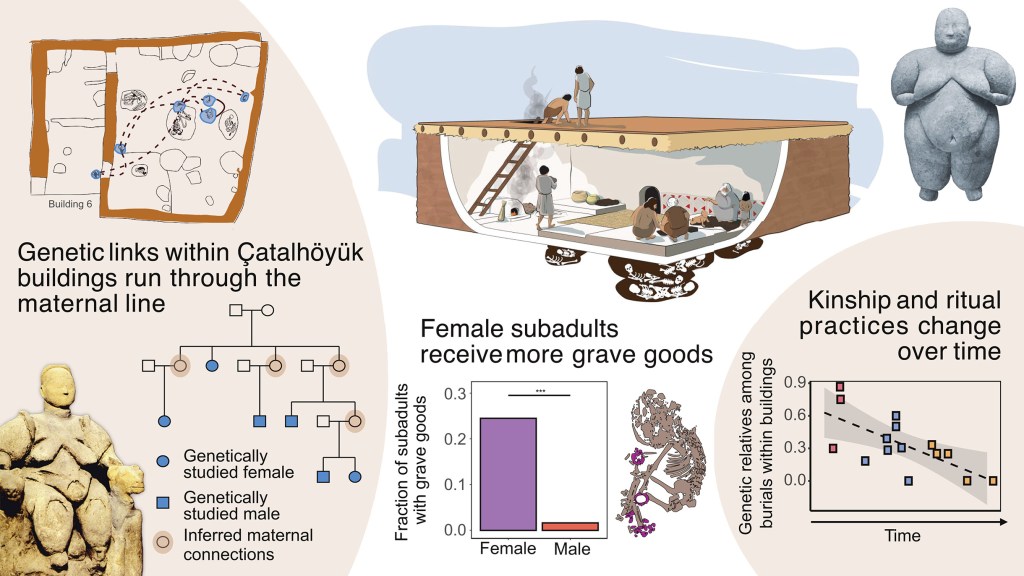
- El asentamiento neolítico de Çatalhöyük (Turquía) sigue aportando novedades interesantísimas. Ocupado durante 1000 años (9000-8000 a. C.), Çatalhöyük es famoso por su tamaño, su aparente estructura social igualitaria y sus enterramientos subterráneos, donde niños y adultos eran enterrados dentro de las casas durante su uso. También es conocido por su gran colección de figurillas femeninas, que se debatieron durante mucho tiempo como posibles representantes de un culto a la «Diosa Madre» e indicios de una sociedad matriarcal. Combinando datos paleogenómicos (de 131 individuos enterrados en 35 casas) con datos arqueológicos y restos óseos de 395 individuos, un nuevo trabajo (Yuncü et al., 2025) concluye que: 1) Entre el 70 % y el 100 % de las veces, las descendientes femeninas permanecieron vinculadas a los edificios, mientras que los varones adultos podrían haberse mudado. 2) Parece existir un trato preferencial para los entierros de niñas y mujeres jóvenes, con cinco veces más ajuares funerarios ofrecidos que a los hombres de su edad. 3) Los patrones de organización social cambiaron con el tiempo: en las primeras fases del asentamiento, los individuos enterrados juntos en casas solían representar a miembros de familias extensas; posteriormente, la composición genética de los enterramientos en casas se volvió menos homogénea, llegándose a encontrar en las últimas etapas grupos de neonatos sin parentesco genético. 4) Que el parentesco genético pudiera haber perdido importancia con el tiempo, junto con la observación de dietas similares en las madres, sugiere la generalización de mecanismos de adopción y la crianza colectiva. Este modelo contrasta con las sociedades contemporáneas a Çatalhöyük en Europa, que tendían a ser patrilocales, y los enterramientos más ricos solían estar reservados a varones. Otro estudio (Koptekin et al., 2025) muestra que las prácticas neolíticas en Anatolia occidental se originaron en interacciones culturales, seguidas de movilidad y mezcla, y, finalmente, la migración a Europa. El análisis comparativo de la similitud cultural y genética en 16 asentamientos neolíticos revela que no existe correlación entre las similitudes culturales y genéticas: las entidades culturales no se corresponden con poblaciones genéticamente homogéneas. Curiosamente, un estudio previo (Mazzucato et al., 2025) sí había encontrado una similitud cultural material entre casas de Çatalhöyük y relaciones biológicas entre individuos.
- Hasta ahora, se pensaba que las islas pequeñas y remotas del Mediterráneo no habían estado pobladas antes de la llegada de la agricultura. A partir de los restos de herramientas de piedra, hogares y fauna cocinada en la cueva de Latnija (Malta), se ha logrado documentar una travesía de más de 100 km en mar abierto, realizada por cazadores-recolectores mesolíticos que viajaron a través del Mediterráneo hace 8,5 ka sin embarcaciones a vela (Bengtsson et al., 2025). Pudieron subsistir en aquellas islas diversificando su dieta, no solo gracias a recursos terrestres, como el ciervo, que no fueron sobreexplotados, sino también aves y alimentos marinos muy variados, incluyendo focas, peces, cangrejos, caracoles, erizos de mar y tortugas.
- Los cazadores-recolectores precerámicos en Colombia, que abarcan un periodo desde hace 6000 años, representan un linaje basal hasta ahora desconocido, que fue reemplazado por poblaciones de Centroamérica asociadas con el complejo cerámico Herrera hace unos 2000 años y sobrevivió durante el período muisca a pesar de importantes cambios culturales. Aquel grupo no dejó contribución genética en las poblaciones posteriores de la región (Krettek et al., 2025).
