
Cerramos un gran año en hallazgos paleoantropológicos. En el último trimestre recopilé 46 publicaciones que ilustran la riqueza acumulada: los cuatro resúmenes trimestrales de 2025 suman 198 novedades. ¡Feliz 2026!
- Combinando modelos tridimensionales de cráneos y métodos comparativos filogenéticos, Gómez-Robles y colaboradores han analizado la velocidad a la que distintas regiones del rostro y del neurocráneo cambiaron a lo largo del tiempo en distintos linajes de primates. El linaje humano duplicó la tasa esperada de cambio morfológico, y esto sucedió en la mayoría de los rasgos craneofaciales, separándose claramente de otros grandes simios y mostrando una selección direccional.
- Al contrario que el desarrollo prolongado en el neurocráneo de Homo sapiens y Homo erectus, en Australopithecus afarensis y Paranthropus robustus los patrones de desarrollo son variables (Braga, Alemseged y Gilissen, 2025). De hecho, Au. afarensis exhibe un crecimiento endocraneal prolongado en comparación con P. robustus, un contraste incluso mayor que el de Pan paniscus y Pan troglodytes, lo que sugiere que el desarrollo endocraneal en Au. afarensis se asemeja más al de H. sapiens que al de P. robustus.
- El análisis 3D morfométrico de las inserciones ligamentarias radiocarpianas (Casado et al., 2025) en 110 radios de homininos fósiles aporta información sobre sus patrones locomotores: Au. afarensis, Au. anamensis y Au. sediba combinaban locomoción bípeda y arbórea similar a chimpancés y orangutanes; P. robustus tenía una locomoción terrestre más similar a la de los gorilas; neandertales y H. sapiens basales muestran una anatomía similar a la de los humanos actuales, pero algo más de robustez orientada a cargas manuales (caza/herramientas).
- El astrágalo ARA-VP-6/500-023 de Ardipithecus ramidus presenta similitudes con los astrágalos de chimpancés y gorilas, adaptados a la escalada vertical y al cuadrupedismo plantígrado terrestre. Además, el fósil tiene rasgos derivados consistentes con un mecanismo de impulso mejorado en el pie de Ar. ramidus. Prang y colaboradores proponen que el último ancestro común de humanos y chimpancés no sería un simio arborícola generalizado, sino un ancestro africano similar a un simio con un repertorio postural que incluía el cuadrupedismo plantígrado terrestre y la escalada vertical, del cual Ar. ramidus representaría un estadio intermedio, con modificaciones incipientes hacia una mayor eficiencia en bipedación. Por otro lado, la bipedación de Ardi sería distinta a la que emplean los grandes simios: su lateral del pie funcionaba mejor para el impulso lateral, dándole un empuje más eficiente al caminar.
- Se han presentado nuevos fósiles de Burtele que permiten atribuir el famoso pie con el hallux oponible encontrado allí en 2009 y otros restos a Australopithecus deyiremeda, coetáneo de Au. afarensis, destacando su mezcla de rasgos arcaicos y derivados, locomoción bípeda aún muy arbórea y dieta basada en plantas C3. Una evidencia de la gran diversidad pliocena en Afar (Haile-Selassie, Schwartz, Prang, et al., 2025).
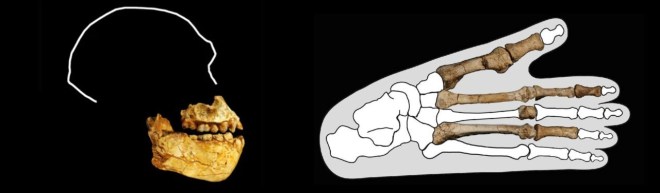
- Martin y colaboradores proponen no atribuir el esqueleto StW 573 («Little Foot») a Australopithecus prometheus, al observar rasgos clave que lo diferencian del holotipo de esta especie, MLD 1, el cual resulta morfológicamente más similar a Au. africanus.
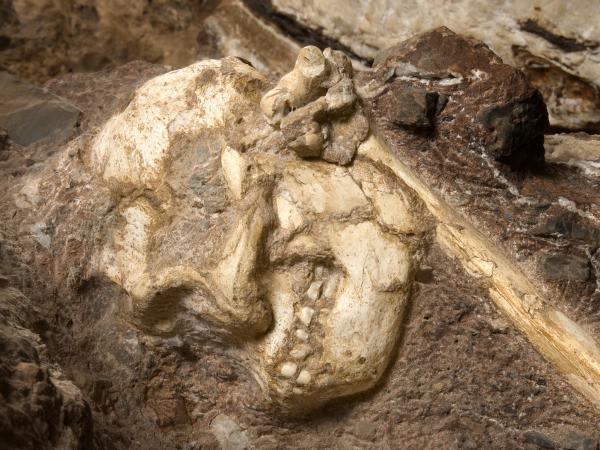
- La reconstrucción virtual a partir de cuatro fragmentos conservados del cráneo DAN5/P1 de Gona (Etiopía), normalmente atribuido a Homo erectus, refleja una bóveda craneal típica de esta especie, pero con un cerebro muy pequeño para su antigüedad (1,5-1,6 Ma) y rasgos faciales y dentales observados más bien en Homo habilis. Este hallazgo respalda la larga persistencia de grupos Homo de cerebro pequeño y plesiomórficos junto con otros grupos que experimentaron encefalización continua durante el Pleistoceno inicial y medio de África (Baab et al., 2025).
- Desde hace un tiempo, el debate «caza vs. carroñeo» no da para mucho más. Los primeros humanos eran omnívoros y obtenían alimento animal mediante la caza o el carroñeo según las condiciones ambientales y sus capacidades tecnológicas y cognitivas. Un nuevo trabajo (Mateos et al., 2025) propone que el carroñeo no debe considerarse una actividad marginal, y supone un recurso abundante, de alta calidad, ubicuo y predecible según la época del año, por tanto, importante para complementar otras formas de obtener alimento cuando era necesario. Las adaptaciones anatómicas, físicas y conductuales permitieron detectar cadáveres a larga distancia, alcanzarlos con relativa rapidez, enfrentarse a otros carroñeros si era necesario, y procesarlos para obtener alimento. Además, también existía carroña procedente de fuentes distintas a la depredación, en particular de megaherbívoros, y podrían haber existido estrategias de reparto de carroña, por ejemplo con los buitres.
- Hace 2,75 Ma, grupos homininos en Namorotukunan (en el lado oriental del lago Turkana, Kenia) dominaban la talla olduvayense para cortar y procesar alimentos, y mantuvieron consistencia técnica y tradición estable de talla durante más de 300 ka, como lo indica la recurrencia tecnológica observada en 1300 lascas en tres capas distintas entre hace 2,75 y 2,44 Ma del mismo yacimiento. Destaca la materia prima seleccionada por su calidad, así como el filo conseguido en muchos artefactos. La inestabilidad del entorno geológico, caracterizado por incendios forestales recurrentes, aridez, desplazamientos de cauces y drásticos cambios ambientales, sugiere una continuidad conductual y cultural (Braun et al., 2025).
- Aunque existen evidencias desde hace 2 Ma en el registro arqueológico de la utilización de artefactos a modo de palillos de dientes, un nuevo trabajo (Towle et al., 2025) subraya otras causas posibles a considerar a la hora de interpretar los surcos observados en la superficie dental.
- Suele hablarse de los bifaces achelenses como herramientas multifuncionales. Y lo eran, pero la multifuncionalidad no era aleatoria ni se expresaba de forma uniforme a lo largo del tiempo. El yacimiento de La Noira (Francia) muestra dos fases ocupacionales achelenses (700 y 450 ka, miles de años) en las que se los bifaces se usaron para diversas tareas, según indican los patrones y la ubicación del desgaste (Rodríguez, Coco, Moncel, et al., 2025). Distintos factores condicionaban la sujección y uso de las herramientas, incluyendo ergonómicos, exigencias funcionales, limitaciones de la calidad de la materia prima, habilidad en la talla y resultados morfológicos imprevistos.
- Durante los últimos 2 Ma, nuestros antepasados han estado expuestos al plomo procedente de actividad volcánica, incendios forestales o procesos geológicos que liberan plomo en agua y alimentos, según indica el análisis de 51 dientes fósiles pertenecientes a especies humanas y prehumanas con antigüedad entre 100 ka y 1,8 Ma. En tejido cerebral cultivado en laboratorio, los organoides que tenían la variante neandertal del gen NOVA1 experimentaron una alteración significativa en la actividad del gen FOXP2, esencial para el lenguaje. La variante humana moderna de NOVA1 pudo ofrecer una resistencia neurológica frente a los efectos tóxicos del plomo, lo que, en un escenario de entornos contaminados naturalmente, habría favorecido una mejor supervivencia y capacidades comunicativas (Joannes-Boyau et al., 2025).
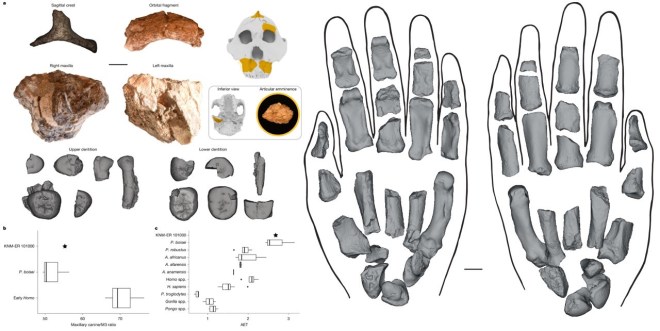
- Se han presentado fragmentos de cráneo, dientes, manos y pies de KNM-ER 101000, atribuido a Paranthropus boisei (Mongle et al., 2025). Su mano muestra notables capacidades para agarrar, únicas entre otros homininos de su época. Compartía con Homo adaptaciones clave para la manipulación y la bipedación. Pero también poseía un agarre poderoso para trepar y procesar alimentos, como las de los gorilas actuales, lo que podría haber condicionado su capacidad para realizar movimientos de precisión.
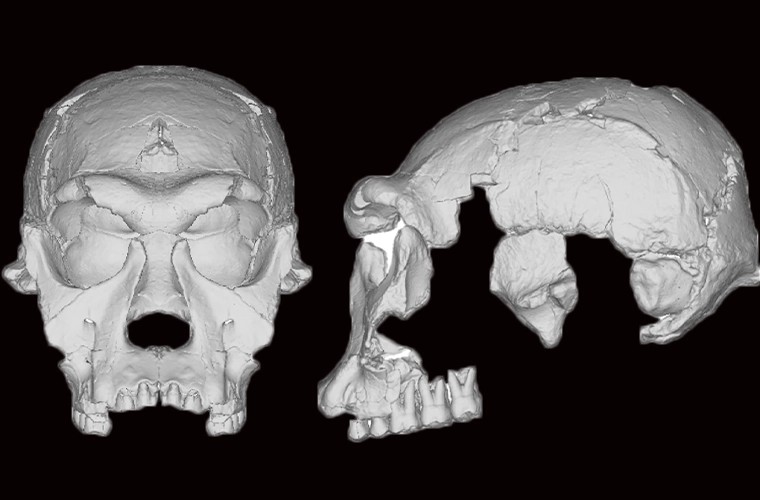
- Nery y colaboradores proponen que las diferencias en las dimensiones coronales de los dientes de Dmanisi respaldan la hipótesis de la coexistencia de dos taxones distintos en este sitio hace 2 Ma, previamente propuestos como Homo georgicus y Homo caucasi, aunque estos son taxones raramente utilizados en la literatura científica. Para ello, comparan afinidades morfológicas y probabilidades posteriores de clasificación de los dientes de tres especímenes (D4500-D2600 o cráneo 5, D2282-D211 o cráneo 2, D2700-D2735 o cráneo 3) con los de 122 homininos fósiles, además de ratios dentales con gorilas y chimpancés para comprobar el dimorfismo. El cráneo 5 se agrupa en el estudio con Au. africanus, mientras que los cráneos 2 y 3 con Homo temprano. Los ratios dentales caen en rangos de grandes simios, lo que permitía asumir un dimorfismo sexual extremo, pero las proporciones coronales, como las del M3 en D4500, sugieren dos taxones: Homo georgicus para el cráneo 5, y Homo caucasi para los otros dos.
- Mediante reconstrucciones cuantitativas de polen y apertura del paisaje en el yacimiento chino de Majuangou (Cuenca de Nihewan, 1,76–1,32 Ma), Fan y colaboradores sugieren que desde el Pleistoceno inferior los homininos se asociaron sistemáticamente con paisajes relativamente abiertos (60–90% de apertura), evitando tanto bosques densos como estepas muy abiertas. Al compilar datos de otros 24 yacimientos pleistocenos de Eurasia y evidencias africanas, proponen que estos paisajes abiertos pero mixtos favorecieron la migración, la caza y la evolución de H. erectus a escala pan‑eurasiática y global.
- La reevaluación del conjunto lítico de Kabwe lo posiciona como una industria transicional ESA/MSA. Debido a la posible asociación con fósiles de Homo heidelbergensis sensu lato del cráneo y restos poscraneales de Kabwe, se sugiere que la etapa inicial de la MSA podría haber incluido a otros miembros de nuestro clado, además de Homo sapiens (Miedzianogora, Shipton y Pope, 2025).
- El estudio de Jakobsson y colaboradores sobre 28 genomas antiguos de individuos procedentes de distintas regiones del sur de África, evidencia una gran diversidad genómica interna, con numerosas variantes exclusivas y una estructura que implica largos periodos de aislamiento relativo, que viene a reforzar propuestas de la configuración de la especie H. sapiens a partir de linajes profundamente diferenciados dentro de África.
- El yacimiento al aire libre de Terra Amata (Niza, sureste francés), datado en MIS 11/10 (~400 ka), fue ocupado de manera estacional y recurrente por grupos achelenses, junto a un delta en un entorno pantanoso. Allí se ha documentado uso del fuego, restos de construcciones y el empleo de algunos materiales recuperados a unos 40 km. García Medrano y colaboradores han analizado su industria lítica, elaborada principalmente con cantos calcáreos locales: herramientas pesadas como choppers, bifaces, hachas de mano y picos, con talla mínima, secuencias centrípetas cortas y lascado simple. Destacan núcleos discoidales elaborados, con incipiente preparación y organización, precursores de la tradición Levallois. Esta flexibilidad tecnológica refleja adaptación a recursos abundantes, con gestión mínima de morfologías naturales, y ejemplifica el umbral tecnológico MIS 12-9 en Europa occidental, con innovaciones en subsistencia, fuego y un mosaico cultural regionalizado.
- Un preprint de Peyrégne y colaboradores describe distintos hallazgos de la secuenciación de alta cobertura del genoma obtenido a partir del esmalte del molar Denisova 25. Este diente perteneció a un individuo masculino de entre 20 y 35 años, que vivió hace unos 200 ka, procedente de la cueva de Denisova. Su ADN muestra que recibió material genético de hibridaciones con neandertales tempranos, de otro linaje superarcaico, es decir, de homininos más primitivos, que dejaron su huella también en los pueblos actuales de Asia y Oceanía, y fue reemplazado por otros grupos denisovanos que, a su vez, se hibridaron con neandertales posteriores.
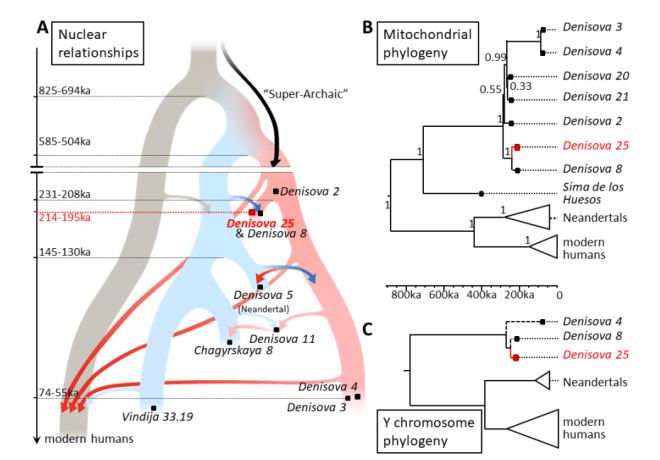
- Los diferentes niveles de ascendencia denisovana en los euroasiáticos se debieron a la mezcla entre diversos linajes humanos modernos tempranos. En particular, los Jomon en el archipiélago japonés y las islas Ryukyu/Okinawa son los que presentan menor ascendencia denisovana entre los individuos de Eurasia oriental, lo que proporciona evidencia de un antiguo linaje de Asia oriental con poca o ninguna ascendencia denisovana. Por el contrario, los primeros humanos modernos del este de Asia continental albergan la mayor ascendencia denisovana en Eurasia oriental, como el individuo de Tianyuan (China, 40 ka, más del 0,2%) incluyendo ascendencia de múltiples grupos denisovanos divergentes, incluso antes del Último Máximo Glacial (Yang et al., 2025).
- Un estudio de Terradillos-Bernal y colaboradores aborda las ocupaciones neandertales en la transición MIS 6–5 (∼140–120 ka) en la cueva de Valdegoba (Burgos, España). Se presenta una nueva falange distal neandertal con rasgos morfológicos clásicos, y un conjunto lítico inédito que confirma la talla musteriense de sílex in situ y una variabilidad tecnológica de los neandertales en la península ibérica durante la transición MIS 6/5, para adaptarse a diversos entornos y recursos empleando diversas estrategias de gestión territorial y de subsistencia.
- Un preprint de Makhro y colaboradores observan variantes genéticas distintas en el gen PIEZO I entre neandertales y H. sapiens. La variante neandertal, similar a la de otros grandes simios, hacía que la hemoglobina retuviera el oxígeno con más fuerza (tal vez como adaptación al frío extremo y la escasez de alimento), mientras que la versión humana moderna permitía liberar el oxígeno de forma más eficiente hacia los tejidos. Cuando la sangre materna tiene demasiado oxígeno ligado a la hemoglobina, menos oxígeno llega al feto a través de la placenta, lo que puede causar hipoxia, restricción del crecimiento fetal o incluso aborto espontáneo. La aparente ventaja en neandertales se habría vuelto perjudicial en los híbridos neandertal-sapiens: las madres heterocigotas portadoras de un alelo neandertal podrían suministrar oxígeno insuficiente a los fetos que heredan dos alelos modernos, lo que reduce su supervivencia. Esta variante está prácticamente ausente en los humanos modernos, lo que correspondería a una fuerte selección negativa.
- La aridificación del paisaje y la competencia entre humanos por recursos escasos habrían provocado el abandono de Homo floresiensis de Liang Bua. Una modelización geoquímica (Gagan et al., 2025) revela una disminución sostenida de la precipitación media anual, de ~1560 a 990 mm, entre hace 76 ka y 61 ka, hasta un mínimo histórico de ~450 mm entre hace 61 ka y 55 ka, junto con una marcada disminución tanto de la abundancia como de la proporción relativa de restos de Stegodon en el registro fósil.
- El ADNmt de un fragmento óseo neandertal (Star 1) datado en 45-46 ka, procedente del refugio rocoso de Starosele, en Crimea, muestra vínculos estrechos con segmentos de ADN de los neandertales de Altái, en Siberia (Pigott et al., 2025). Grupos neandertales del este de Europa viajaron unos 3000 kilómetros hacia Asia, donde dejaron un legado genético y también cultural vinculado a la industria micoquiense, un desarrollo final del Achelense. Los desplazamientos ocurrirían durante períodos cálidos que, según estudios geológicos, se ubican en dos momentos: entre 120-100 ka y 60 ka. Por otra parte, la abundancia de fósiles de caballos en Starosele indica que formaban una parte importante de su alimentación.
- Un estudio de los restos neandertales de 41-45 ka de la cueva Troisième de Goyet, Bélgica (Cosnefroy, Crevecoeur, Semal, et al., 2025), que combina paleogenética, isótopos, morfología ósea y biomecánica, indica que al menos seis individuos fueron canibalizados. Sus huesos presentan fracturas en fresco, marcas de corte, impactos de percusión y evidencias de uso como herramientas óseas. Fueron cuatro mujeres adolescentes o adultas jóvenes, un niño de entre 6 y 12 años y un neonato. Las tibias de las mujeres muestran secciones casi circulares, asociadas a cargas multidireccionales moderadas, no a desplazamientos largos o repetitivos, pero los análisis isotópicos de azufre indican que las mujeres procedían de otra región, y tenían dietas similares entre sí. Por otra parte, tenían baja estatura (143-156 cm) y eran gráciles, con diámetros corticales reducidos. Los autores proponen un posible contexto de conflictos territoriales o tensiones entre comunidades neandertales, en que ocurriría la predación selectiva de individuos vulnerables (mujeres jóvenes y niños), exocanibalismo (consumo simbólico o violento de individuos de otros grupos), ataques destinados a reducir la fertilidad de grupos rivales, y secuestro o captura de individuos.
- Se ha presentado un útil óseo procedente del Abri du Maras (Ardèche, Francia), fechado en el MIS 5 (130-80 ka), interpretado como herramienta neandertal para desollar o despellejar carcasas. Los análisis traceológicos y experimentales indican que las huellas de uso parecen deberse al raspado y despegue de piel para fines funcionales como vestimenta o coberturas (Doyon et al., 2025).

- El yacimiento de Barnham, en Suffolk (Reino Unido), ha proporcionado el hallazgo más antiguo y mejor documentado de producción de fuego por grupos humanos (Davis et al., 2025). Se trata de hogares fechados en la primera mitad del Hoxniano (MIS 11c, ~427-415 ka, miles de años) que documentan un uso y mantenimiento recurrente del fuego, asociados a herramientas calentadas y, de manera destacada, a dos fragmentos de pirita con huellas microscópicas de uso
- Tres artefactos de ocre procedentes del yacimiento micoquiense de Zaskalnaya (Crimea) presentan características que van más allá del uso utilitario: uno es similar a un lápiz con afilados repetidos, otra parece ser un fragmento de lápiz y una tercera presenta superficies grabadas y pulidas. Estos rasgos sugieren la producción intencional de marcas y un uso selectivo. Aunque aplicaciones prácticas son plausibles, como el procesamiento de pieles, d’Errico y colaboradores proponen el uso simbólico, basándose en comportamientos simbólicos de otros neandertales en la región.
- La capacidad de procesar una amplia gama de alimentos vegetales representó un umbral clave en la evolución humana ya desde los primeros grupos humanos, más que un paso en el camino hacia la agricultura. Evidencias arqueológicas en yacimientos en todo el mundo sugieren que distintas comunidades molían semillas silvestres, machacaban y cocinaban tubérculos ricos en almidón y desintoxicaban frutos secos amargos muchos milenios antes del desarrollo de la agricultura (Florin y Ramsey, 2025).

- En otro trabajo, se concluye también que neandertales y humanos modernos transformaron los ecosistemas europeos miles de años antes del surgimiento de la agricultura, influyendo en los patrones de vegetación que caracterizaron el continente (Nikulina et al., 2025). Comparando simulaciones informáticas de modelos climáticos y el registro paleopalinológico, incluso añadiendo factores como incendios naturales, pastoreo y cambios de temperatura, los resultados no coinciden sin sumar la intervención humana. Con ella, los modelos indican que las actividades humanas alteraron de manera significativa la composición y apertura de los paisajes: el uso del fuego promovió la expansión de pastizales y la apertura de bosques, mientras que la caza redujo la presión de pastoreo, favoreciendo el crecimiento de zonas más densas en arbustos y árboles. Los cazadores-recolectores del Mesolítico habrían influido en el 47% de la distribución de los tipos de plantas, y los neandertales en cerca del 6% de la vegetación y el 14% del grado de apertura del entorno.
- Buena parte del debate sobre las adaptaciones respiratorias de Homo neanderthalensis al clima frío tradicionalmente ha girado en torno a ciertos rasgos de la cavidad nasal interna, considerados tradicionalmente autapomorfías de esta especie: una cresta medial vertical en la pared nasal interna, un engrosamiento medial adyacente y la supuesta ausencia de un techo óseo sobre el canal nasolagrimal. La limitación y fragmentación del registro fósil habían dificultado la verificación de estos rasgos, pero la nueva evidencia en el neandertal de Altamura permite descartarlos de manera sólida, aportando nueva información sobre la morfología nasal neandertal y su significado adaptativo (Buzi et al., 2025).

- Un estudio de Amadei, Lin y Fattorini aporta un marco cuantitativo para evaluar la hipótesis de la desaparición del linaje neandertal por un proceso impulsado, al menos en parte, por el flujo génico desde poblaciones mayores de Homo sapiens. No excluye otras explicaciones, pero demuestra que la mezcla genética recurrente por sí sola es un mecanismo teóricamente suficiente para explicar la pérdida del linaje neandertal en el largo plazo.
- Un modelo de Shao et al. (2025) indica que los neandertales ibéricos, fragmentados en pequeñas poblaciones costeras y demográficamente debilitados por el estrés climático del MIS 3, persistieron hasta hace unos 41-38 ka, coincidiendo parcialmente con la llegada de los humanos modernos al noroeste peninsular. Estos últimos, con mayor conectividad y resiliencia, se expandieron rápidamente por rutas atlánticas e interiores, sustituyendo gradualmente a los neandertales en un proceso de solapamiento y reemplazo demográfico más que de extinción súbita.
- El estudio de Uttley y colaboradores señala algunas diferencias mínimas en el ADN que pudieron contribuir a la mandíbula más robusta de los neandertales y ayudarían a entender la variación facial humana. Identificaron un pequeño segmento del genoma neandertal, con solo tres cambios respecto al humano, que regula la activación del gen SOX9, crucial para el desarrollo facial. Al introducir las versiones humana y neandertal en embriones de pez cebra, ambas se activaron en células formadoras de la mandíbula, pero la variante neandertal mostró una actividad significativamente mayor. Esa mayor activación incrementa el efecto de SOX9 y favorece un crecimiento mandibular más amplio.
- Se ha presentado un incisivo superior central hallado en Gua Dagang («Cueva del Comerciante»), norte de Borneo (Malasia), datado en 52-55 ka, cuya morfología no encaja con H. sapiens (Curnoe et al., 2025). Hasta ahora solo existía registro arqueológico atribuido a H. sapiens en torno a 46–50 ka en la cercana Gran Cueva de Niah. Rasgos que se apartan del rango de H. sapiens y se solapan con H. erectus s.l., H. neanderthalensis y otros homininos del Pleistoceno medio son: corona muy ancha, relativamente corta labiolingualmente, notable forma de pala, esmalte labial muy convexo y compleja morfología en la unión esmalte‑dentina.
- Los primeros cambios culturales notables en el sur de China surgieron hace unos 45 ka, con la significativa proliferación de yacimientos que reflejan tecnologías complejas y objetos simbólicos observados en otras partes de Eurasia. Zhou y colaboradores proponen que los abruptos cambios culturales en este periodo se deben a la llegada y dispersión de humanos modernos entre hace 40 ka y 50 ka, como parte de una migración global más amplia durante el Pleistoceno final.
- Distintos conjuntos líticos contemporáneos del Ahmarense en el Levante mediterráneo y del Protoauriñaciense europeo hace unos 42 ka sugieren que el desarrollo europeo no fue una influencia levantina, sino un proceso de innovación independiente (Falcucci y Kuhn, 2025). El foco del estudio fue principalmente las láminas fabricadas en los sitios de Ksar Akil en Líbano, y Grotta di Fumane, Riparo Bombrini y Grotta di Castelcivita en Italia. Aunque las dos industrias se caracterizan por el predominio de la tecnología laminar, los detalles de las tecnologías empleadas, desde la forma en que se explotaban los núcleos y los tipos de percusión utilizados hasta los productos (láminas frente a microláminas), son altamente divergentes. Los autores plantean interesantes reflexiones sobre los orígenes de los dos tecnocomplejos, así como del chatelperroniense.
- Groucutt y colaboradores publican importantes evidencias arqueológicas y paleontológicas en el noreste árido de Arabia, hasta ahora poco explorado. En una de las muchas cuevas, Murrubeh, hallaron miles de huesos de distintas especies acumulados por hienas y procesos naturales, junto con registros de microfauna que ayudan a reconstruir paleopaisajes. Identificaron herramientas de sílex desde el Paleolítico inferior y abundantes del Paleolítico medio (200-50 ka en Arabia). Mientras que en el noroeste de Arabia existen miles de estructuras megalíticas del Holoceno, en la zona del estudio solo han encontrado una estructura reseñable, posiblemente un túmulo de la Edad del Bronce. Los resultados muestran una presencia humana discontinua, vinculada a antiguos recursos hídricos como acuíferos, y abundantes cuevas, de las que algunas de ellas aportan un registro climático de ocho millones de años.
- Gandini y colaborares concluyen que la colonización de Sahul ocurrió hace unos 60 ka, apoyando la hipótesis de la «cronología larga» para este proceso y proponiendo al menos dos rutas de llegada distintas, norte y sur. Llegan a esa conclusión analizando 2456 mitogenomas de poblaciones indígenas de Australia, Nueva Guinea y Oceanía, combinando la reevaluación de la tasa de mutación mitogenómica con datos genómicos, del cromosoma Y, arqueológicos y climatológicos. Las divergencias genéticas clave se corresponden mejor con una llegada temprana (~60 ka). Además, la amplitud y distribución de linajes mitocondriales se explican mejor mediante asentamientos iniciales más antiguos y múltiples rutas de entrada.
- El Puente Terrestre de Bering probablemente no surgió hasta hace unos 35 ka, según un nuevo estudio (Wanket et al., 2025). Previamente, se estimaba que podría estado inundado entre hace 46 y 35,7 ka (Farmer et al., 2022).
- Se ha presentado un nuevo enterramiento paleolítico: Romito 9, en el sur de Italia, es el cuerpo de una niña de 11–12 años fechado en unos 19 ka. Su cuerpo en posición flexionada está acompañado por ocre y adornos (destacando 99 caninos de ciervo y 1462 conchas perforadas de moluscos marinos), indicando una continuidad cultural en la transición al Paleolítico superior final en la región (Martini et al., 2026).
- En Mezhyrich (Ucrania), hace unos 18 ka, un grupo humano construyó al menos cuatro estructuras circulares hechas con huesos de mamut, de entre 12 y 24 m2. Alrededor de ellas se hallaron pozos con herramientas y adornos, además de áreas delimitadas que habrían servido como talleres o espacios de trabajo. Según una cronología radiocarbónica revisada, estos conjuntos se utilizaron durante menos de 429 años (Chu et al., 2025).
- Se han hallado en Norteamérica distintos conjuntos líticos de unos 13,5 ka con tecnologías duales de núcleo-hoja y bifaz, similares a las del Paleolítico superior del noreste de Asia, y que aparecieron en Japón hace unos 20 ka. Madsen y colaboradores proponen que un grupo aislado en la región Paleo-Sajalín-Hokkaido-Kuriles migró hacia el sur de Beringia y la costa noroeste americana, entre hace 22 y 18 ka, dispersándose ampliamente hace 15 ka.
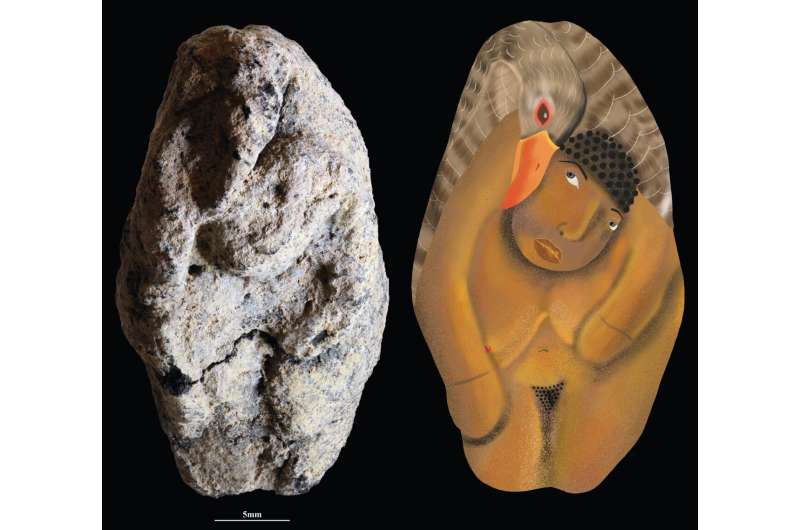
- Una figura femenina con un ganso hallada en Nahal Ein Gev II (Israel), datada en unos 12 ka, constituye una de las representaciones simbólicas más tempranas de la cultura natufiense (15-11 ka, últimos cazadores-recolectores en fase temprana hacia el sedentarismo). Mide 3,7 cm, fue modelada en arcilla y endurecida mediante cocción controlada. Los análisis revelan restos de ocre rojo y la huella dactilar de quien la elaboró. Representa una escena interpretada como un relato mítico: el ganso es un animal consumido pero también cargado de significado ritual, y aparece vivo y rodeando a la mujer en una expresión animista. El uso de volúmenes, luces y sombras indica un manejo precoz de recursos visuales complejos que se consolidarían en el Neolítico. La pieza se encontró en un contexto de de enterramientos y depósitos ceremoniales, reforzando su carácter ritual y simbólico (Davin et al., 2025).
- Hallazgos arqueológicos en distintas localizaciones del interior de la Meseta central ibérica refuerzan la hipótesis de que la ocupación humana del Pleistoceno final en la región fue más frecuente de lo que se suponía tradicionalmente (Aragoncillo et al., 2025).
- El estudio de 30 trozos de brea de corteza de abedul, incluyendo tacos masticados, herramientas con mango y cerámica reparada, ofrece perspectivas sobre la vida de agricultores de hace 6000 años en 9 yacimientos arqueológicos de los Alpes y sus alrededores (White et al., 2025). Se consiguió extraer ADN de 19 muestras y determinar el sexo biológico de 16. Algunos parecen haber sido masticados por varias personas. Los 4 trozos usados para enmangar herramientas de piedra contenían ADN masculino, mientras que los 3 usados para reparar cerámica contenían ADN femenino. Aunque la muestra es pequeña, el trabajo abre camino para nuevos estudios sobre la división del trabajo en comunidades antiguas. Por otra parte, en los trozos masticados se halló ADN de trigo, cebada, avellano y haya, probables restos de alimentos. En la cerámica se halló ADN de guisantes, avellanos y ovejas, probables alimentos almacenados. En puntas de flecha se halló ADN de pescado y jabalí, y ADN de pino en las herramientas con mango, lo que sugiere que se pudo haber añadido resina de pino para hacer el alquitrán más pegajoso. Los trozos masticados también conservaron restos de saliva antigua, con microbioma oral más parecido al de las bocas de humanos modernos que a los presentes en el cálculo dental.
- Comparando en experimentos controlados 191 disparos con 60 puntas óseas montadas en lanzas y flechas, Kitagawa y colaboradores sugieren que las puntas óseas auriñacienses pueden funcionar eficazmente como armaduras de lanza o de flecha, con profundidades de penetración letales en ambos casos. Los patrones de fractura dependen sobre todo del material (asta frente a hueso) y del tamaño de la punta, no del sistema de propulsión. Por ello, proponen que no es posible distinguir de forma fiable entre lanza y flecha solo a partir de la morfometría y las huellas de impacto de las puntas, y sugieren el uso de estrategias de caza con útiles diversos desde el inicio del Paleolítico superior en Eurasia.
- Se ha identificado un linaje prehistórico hasta ahora desconocido en el centro de Argentina, mediante el análisis de ADN antiguo proveniente de 238 individuos indígenas de hasta 10 ka de antigüedad. Su comparativa con otros 588 genomas antiguos de toda América, indica que este linaje habría emergido hace unos 8500 años y persistió como componente mayoritario de la ascendencia en el centro de Argentina. A pesar de la proximidad geográfica con otras poblaciones, los individuos de este linaje parecen haberse mezclado poco con grupos externos, aún cuando existieron cambios climáticos, transiciones económicas (por ejemplo hacia la agricultura) y migraciones de otras poblaciones. Sin embargo, su herencia genética estable difiere de la diversidad cultural y lingüística este grupo antiguo (Maravall-López et al., 2025). Estudios previos habían mostrado la evidencia más antigua de presencia humana en la región en Arroyo Seco, en la Pampa argentina, de unos 14 ka, y una diferenciación hace 9 ka en tres linajes: uno en los Andes centrales, otro en las tierras bajas tropicales de la Amazonia y un tercero al sur, en las Pampas, Chile y la Patagonia.

Pingback: Websites via Bluesky 2026-01-01 – Ingram Braun