Science communication of human evolution faces challenges derived from the complexity of paleoanthropological evidence, the provisional nature of scientific interpretation, and the persistence of cultural misconceptions such as linear models of evolution. This article examines the epistemological, cultural, and communicative factors that shape public understanding of human evolution and analyzes the limitations of traditional deficit-based communication models.
Using a critical review of science communication literature and illustrative cases from paleoanthropology, the study proposes strategic approaches that balance scientific rigor, explicit representation of uncertainty, and accessibility for diverse audiences.
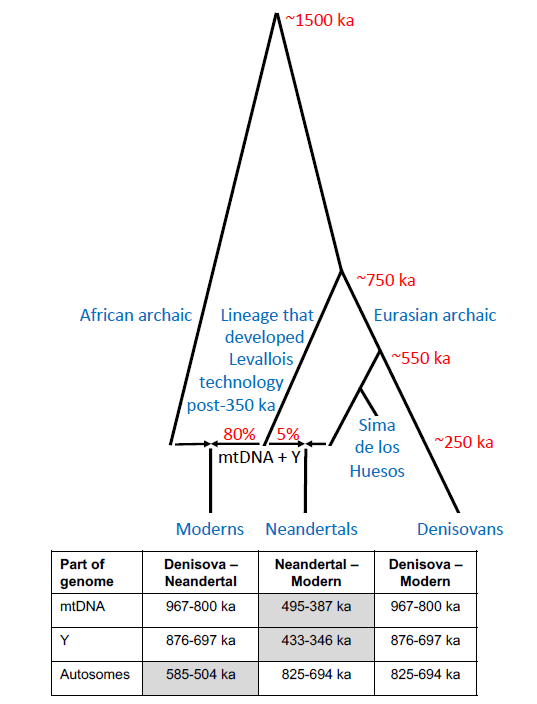
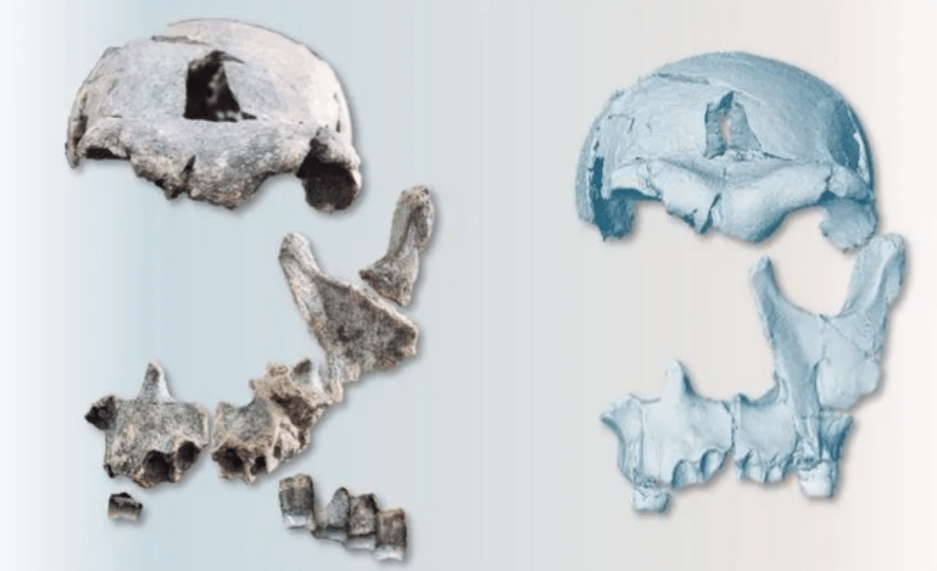
Within this framework, Digital Paleoanthropology is conceptualized as an emerging paradigm that integrates scientific visualization, controlled narrative, and digital participation to enhance the understanding, engagement with, and appreciation of paleoanthropological knowledge. Case examples such as Lucy and Homo naledi demonstrate how visual representation, early knowledge dissemination, and digital co-creation can reshape public interpretive frameworks around human origins.
The article concludes that effective communication of human evolution requires integrating conceptual clarity, epistemic transparency, narrative design, and participatory practices, positioning science communication as a collaborative process that strengthens public understanding of evolutionary science.
Sáez, R. (2026). La comunicación científica de la evolución humana: del déficit al diálogo digital. SciComm Report, 6, 1–21. https://doi.org/10.32457/scr.v6.3468