
< English version below>
Nuestro conocimiento sobre la evolución de los homininos más antiguos se basa principalmente en dos regiones africanas: los ricos yacimientos del Gran Valle del Rift en el este, y los sistemas kársticos del sur.
Sin embargo, en las últimas dos décadas varios ejemplos notables han roto esa norma, y tenemos algunos hallazgos importantes también en el centro de África. Destaca Sahelanthropus tchadensis, sobre el que escribí en este otro artículo: Toumaï, esperanza de vida. Hoy me detengo en un pequeño conjunto de fósiles definido por sus descubridores como Australopithecus bahrelgazali.
Un poco de historia
Ya en 1961, el Chad despertó la atención de los paleontólogos con el hallazgo de un fósil de homínido definido como Tchadanthropus uxoris, que finalmente resultó ser un cráneo erosionado de Homo sapiens. En 1994 una expedición francesa dirigida por Michel Brunet volvió a realizar trabajos paleontológicos en el desierto del Chad, con el reconocimiento de 11 yacimientos con mamíferos de entre 3-4 Ma al este de Koro Toro, pero sin homininos.
En 1995 el mismo equipo encontró cerca del río Bahr el Ghazal, a menos de 1 km de aquellos yacimientos, restos de fauna que habían quedado descubiertos tras las lluvias del año anterior, y también por fin el deseado hominino. Se trataba tan solo de una mandíbula, identificada como KT12/H1, que conserva siete dientes (un incisivo, dos premolares y dos caninos). En 1996 apareció otro premolar, y en 2000 otro fragmento de mandíbula. La antigüedad de los restos se estimó en 3-3,5 Ma mediante la asociación con la fauna del mismo yacimiento.
Ese fósil fue apodado ‘Abel’ en homenaje a Abel Brillanceau, geólogo amigo de Brunet.
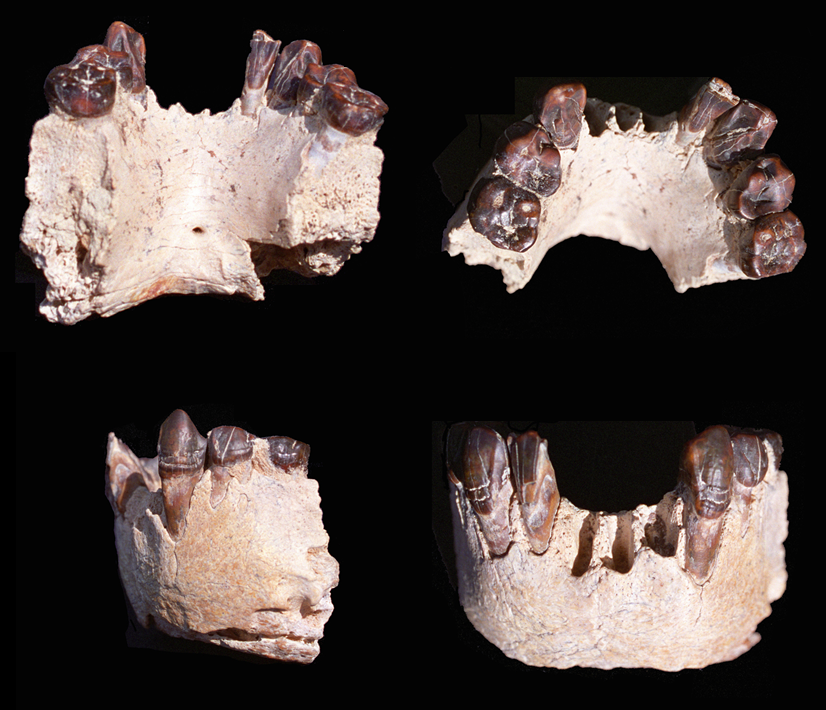
Australopithecus bahrelgazali KT12/H1. Photo credit: Alain Beauvilain
¿Por qué una especie diferente?
Estos son los principales rasgos que llevaron a sus descubridores a proponer una nueva especie para estos fósiles: Australopithecus bahrelgazali, distinta a la de sus contemporáneos del este, Au. afarensis: Sigue leyendo





