
- La selección de herramientas de piedra por parte de los chimpancés salvajes para romper nueces en Bossou (Guinea), en experimentos controlados que introducen rocas desconocidas para esta población, podría reflejar las técnicas de los homininos que desarrollaron la industria olduvayense (Braun et al., 2025). Los chimpancés seleccionan yunques y martillos en función de las propiedades mecánicas, características no directamente visibles para el individuo: piedras más duras para los martillos y más blandas para los yunques, lo que indica que comprenden propiedades específicas para funciones distintas. Los adultos identifican las propiedades mecánicas mediante el aprendizaje individual, y los jóvenes a menudo reutilizan las herramientas seleccionadas por los adultos.
- El origen de la talla lítica pudo haber sido un proceso emulativo por parte de los homininos que buscaban reproducir objetos afilados producidos de forma natural («naturalitos») y que ya eran muy demandados, y no tanto la creación intencional o por accidente de lascas afiladas y la posterior aplicación a objetos para cortar (Eren et al., 2025).

Ejemplos de objetos de piedra con filos producidos naturalmente. Crédito: Michelle R. Bebber, Metin I. Eren y Alastair Key
- La orientación acetabular de los Australopithecus era esencialmente similar a la humana hace 3,6 Ma, atendiendo a la comparativa de los acetábulos en una muestra de primates antropoides existentes y homininos fósiles que incluyen Australopithecus afarensis (AL 288–1, KSD-VP-1/1), Australopithecus africanus (Sts 14), Australopithecus sediba (MH2) y Homo neanderthalensis (Kebara 2). El cambio en la orientación acetabular de los homininos fue probablemente un rasgo seleccionado para la locomoción (Lawrence, Hammond, Ward, 2025).
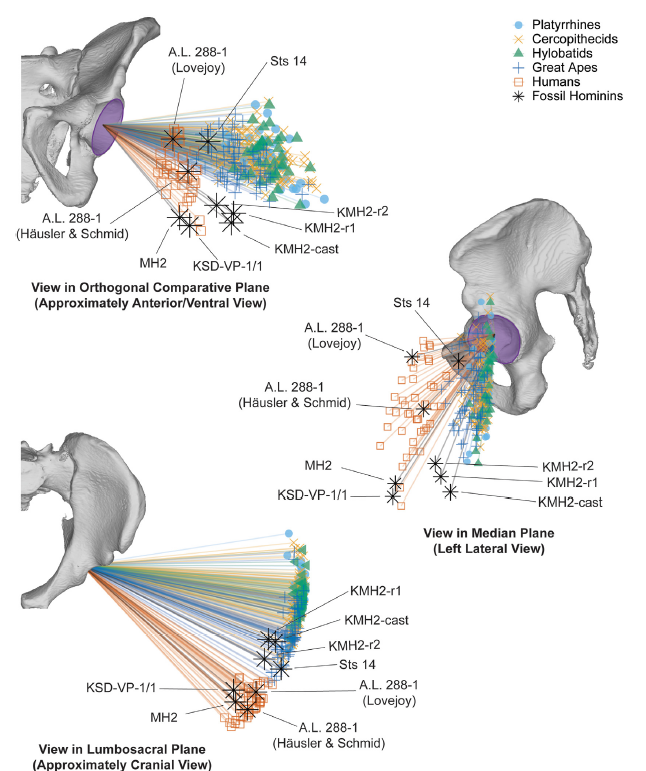
Vectores de orientación acetabular en 3D (vectores normales al plano acetabular) superpuestos en una pelvis humana y mostrados en vistas que representan el plano medio, el plano lumbosacro y el plano comparativo ortogonal. Figura 4 en A.B. Lawrence, A. S. Hammond, C. V. Ward (2025)
- Se han comparado los isótopos de carbono y nitrógeno unidos al esmalte dental de los dientes fósiles de 43 mamíferos de hace 3,5 Ma de Sterkfontein (Sudáfrica), incluidos 7 individuos diferentes de Au. africanus, con los dientes de mamíferos africanos modernos (Lüdecke et al., 2025). Cuando los animales digieren los alimentos, sus cuerpos tienden a utilizar y eliminar por la orina, las heces y el sudor más el isótopo ligero de nitrógeno (14N) que el isótopo pesado (15N), el cual queda más en sus cuerpos en comparación con los alimentos que comen. Las plantas contienen mucho 14N pero no mucho 15N; los herbívoros tienen más 15N y menos 14N en sus cuerpos que las plantas que comen; y los carnívoros tienen mucho 15N y no mucho 14N. El estudio observa que los australopitecos parecen haber tenido una dieta variada, pero no rica en carne de mamíferos. Este hallazgo podría implicar que las habilidades de los australopitecos para adaptarse a ecosistemas de sabana pudo haber sido anterior al cambio a una dieta rica en carne que potenció la evolución del cerebro. Tampoco se puede descartar una dieta rica en termitas, alimento que contiene relativamente menos nitrógeno del tipo que se encuentra en la carne de los mamíferos.
- Se ha publicado el conjunto articulado del hueso coxal, el fémur casi completo y la tibia completa de un adulto joven atribuido a Paranthropus robustus (Pickering et al., 2025), procedente de Swartkrans Miembro 1, cuya datación es 1,7-2,3 Ma. Este fósil, SWT1/HR-2, es uno de los homininos adultos más pequeños conocidos en el registro fósil, probablemente una hembra con una estatura de poco más de un metro y un peso de unos 27 kg. El borde del hueso coxal estaba mordido por un leopardo.
- Estudiando el proteoma de un molar se ha logrado determinar el sexo (masculino) del espécimen Sts 63 de Australopithecus africanus, procedente del Miembro 4 de Sterkfontein. Un prometedor trabajo para seguir explorando el potencial de los estudios paleoproteómicos (Madupe et al., 2025).
- Al mismo tiempo que grupos de Homo erectus comienzan a desarrollarse en África, con los ejemplares más antiguos conocidos datados en 2 Ma, otros homininos (¿también de la misma especie?) en Europa hace 1,95 Ma procesaban huesos de animales dejando marcas de corte, en la localidad de Grăunceanu, en el valle del río Olteț, sur de Rumanía (Curran, S.C., Drăgușin, V., Pobiner, B. et al., 2025).

Especímenes con marcas de corte de hace 1,95 Ma hallados en Grăunceanu. Crédito: Curran, S.C., Drăgușin, V., Pobiner, B. et al. (2025)
- Un nuevo trabajo (Méndez-Quintas, Domínguez y Mussi, 2025) analiza el origen del Achelense a partir del amplio conjunto lítico del sitio de Gombore I nivel B (Melka Kunture, Etiopía). Destaca que esta industria conserva muchas características típicas de la tecnología olduvayense, como el uso extensivo de métodos de núcleo y lasca, pero también otras innovaciones que van más allá de la producción de lascas de gran tamaño o la talla de bifaces, como la segmentación espacial de las secuencias operativas, la mayor complejidad en la selección de materias primas y el retoque de herramientas, así como la aparición de sistemas complejos de producción de lascas, como la Kombewa y la Tecnología de Núcleo Preparado.
- Un importante hallazgo sitúa el origen de la producción sistemática de útiles óseos hace 1,5 Ma en la Garganta de Olduvai (de la Torre, Doyon, Benito-Calvo, et al., 2025), un millón de años antes de lo que se conocía. Se trata de 27 herramientas elaboradas a partir de huesos de hipopótamo (8), elefante (6), bóvidos (2) y de otros animales no identificados. Se comenzaron a descubrir en 2015 en el complejo T69 del barranco occidental del yacimiento FLK (Frida Leakey Korongo). Los fabricantes adaptaron artefactos líticos para manipular los huesos, extrayendo lascas y dando la forma deseada a la herramienta.
- Se han documentado esferas de roca en 8 yacimientos achelenses de Melka Kunture, en los alrededores volcánicos del Alto Awash, que abarcan más de un millón de años, desde Gombore IB (al menos 1,7 Ma) hasta Garba I (0,6 Ma). A diferencia de las encontradas en otros sitios pleistocénicos (como los 150 artefactos de ‘Ubeidiya, 1.4 Ma, pulidos para generar la forma redondeada), estas esferas no son el producto final o subproducto de la actividad antrópica, sino formas naturales de origen volcánico cuidadosamente seleccionadas y recogidas, tanto por Homo erectus como por especies sucesoras, en afloramientos o depósitos secundarios que variaron a lo largo del tiempo (Mussi, 2025). Se trata de una curiosa colección deliberada de estos artefactos, por algún motivo. Es probable que las marcas de golpes y los planos de superficie desgastados sean el resultado de actividades de percusión. A. Muller et al., (2023) levantaron el siguiente mapa con los sitios conocidos con esferoides documentados.
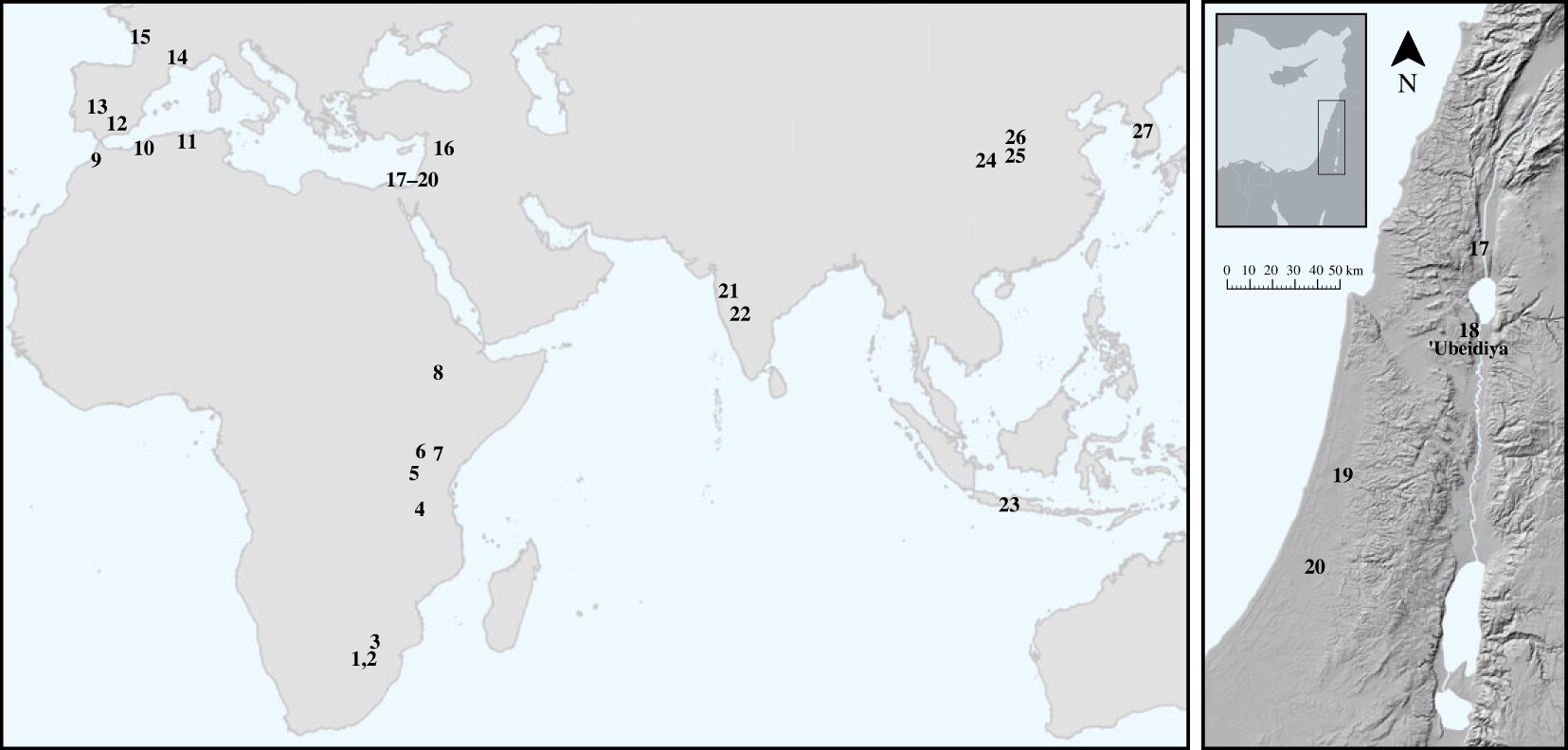
Mapa con artefactos antrópicos redondeados y esferoides. 1. Sterkfontein; 2. Kromdraai; 3. Cave of Hearths; 4. Isimila; 5. Olduvai Gorge; 6. Olorgesailie; 7. Isenya; 8. Melka Kunture; 9. Thomas Quarry I; 10. Ternifine; 11. Ain Hanech, El Kherba and Ain Boucherit; 12. Barranco León; 13. Santa Ana; 14. Bois-de-Riquet US4; 15. La Quina; 16. Hummal; 17. North of Bridge Acheulean locality of Gesher Benot Ya’akov; 18. ‘Ubeidiya; 19. Qesem Cave; 20. Revadim; 21. Chikri-on-Pravara; 22. Hungsi Valley; 23. Ngebung; 24. Lantian; 25. Sanmenxia; 26. Fenhe River sites; 27. South Korean river basin sites. Crédito: Antoine Muller et al (2023)
- Continuando con los esferoides, estudios experimentales (Assaf et al., 2025) muestran cómo los homininos del Paleolítico inferior usaban estos artefactos para extraer médula ósea y, en esta acción, la destreza técnica era clave. Se requiere planificación, conocimiento de la anatomía ósea y del uso de herramientas, así como habilidad y experiencia. La postura corporal, gestos, agarre y manejo de las herramientas influyen en la intensidad y la dispersión de los rastros de desgaste en las herramientas.
- También a través de un estudio experimental se han tratado de comprender (Paixão et al., 2025) los criterios de toma de decisiones involucrados en el uso de materias primas hace 1,6 Ma por parte de los fabricantes de herramientas del Achelense inicial en Melka Wakena, en las tierras altas de Etiopía. El trabajo concluye que los homininos evaluaron múltiples factores a la hora de seleccionar, adquirir y usar diversas materias primas, no sólo la idoneidad para producir y diseñar herramientas, sino también la eficiencia y durabilidad de los materiales.
- Se ha publicado la cara parcial humana más antigua de Europa occidental, procedente de la Sima del Elefante (Atapuerca, España), con una antigüedad de 1,1-1,4 Ma (Huguet, Rodríguez-Álvarez, Martinón-Torres et al., 2025). Se trata del fósil ATE7-1, apodado Pink.

Fósil humano ATE7-1 (Pink). Crédito: María D. Guillén / Elena Santos / IPHES / CENIEH
- Hace 1,2 Ma, grupos de Homo erectus se adaptaron a un entorno dominado por matorrales semidesérticos en Engaji Nanyori (Garganta de Olduvai, Tanzania), reconstruido a partir de distintas disciplinas por Mercader, Akuku, Boivin y otros (2025). A pesar de estos desafíos, el Homo erectus desarrolló herramientas especializadas, ocupó repetidamente paisajes fluviales, regresando repetidamente a ríos y fuentes de agua concretas a lo largo de milenios. Este trabajo recupera el carácter de hominino generalista de Homo erectus, que habitaría paisajes complicados adaptándose con recursos limitados, una habilidad clave para su adaptación y dispersión dentro y fuera de África, llegando hasta Arabia y Eurasia.
- Los casos más antiguos conocidos de carnicería y consumo humano de mamíferos carnívoros corresponden a una zorra y un oso hallados en el nivel TD6 del yacimiento de Gran Dolina (Atapuerca, España, 850 ka), asociados a Homo antecessor, aumentando en cronologías posteriores dicho consumo (López-Parés y Cáceres, 2025). Este hallazgo se deriva del análisis de todos los casos documentados hasta ahora de consumo humano de mamíferos carnívoros durante el Paleolítico y el Epipaleolítico/Mesolítico en la península ibérica. El trabajo también propone interpretaciones sobre las posibles causas de este consumo, vinculadas a alimentación de emergencia, aprovechamiento multifuncional de recursos, conflictividad interespecífica o motivaciones simbólicas y rituales. Asimismo, se reflexiona sobre el papel de la caza y el consumo de mamíferos carnívoros en la evolución humana, destacando su relación con la transición de los homininos al rol de grandes depredadores, la aparición de nuevos patrones alimentarios y la extinción de los grandes mamíferos carnívoros en Europa.
- Se han encontrado diversos taxones vegetales a partir de granos de almidón en herramientas de percusión (yunques y percutores) hechas en basalto hace unos 780 ka en Gesher Benot Ya’aqov, al sur del valle de Hula (norte de Israel). Se trata de alimentos que proceden de diferentes nichos ecológicos, estacionalidad y modos de recolección y procesamiento, como bellotas, granos de hierba, castañas de agua, rizomas de nenúfar amarillo y semillas de leguminosas. Los autores destacan la difícil preparación de estos alimentos para su consumo en una antigüedad tan grande, ya que requeriría importantes habilidades cognitivas, largo tiempo de elaboración y herramientas específicas (Ahituv et al., 2025).

Yunque (izquierda) y percutor (derecha) utilizados para procesar plantas. Crédito: equipo de Gesher Benot Ya’akov
- La reconstrucción de una extremidad inferior de Homo naledi proporciona nuevos datos sobre el esqueleto poscraneal de esta especie, destacando el pequeño tamaño de las superficies articulares y los diámetros corticales de los huesos largos. Sus piernas y pies eran muy similares a los de los seres humanos actuales, lo que sugiere que caminaba de manera similar. Sin embargo, los rasgos señalados, junto con su baja masa corporal y una pelvis relativamente más ancha en comparación con la nuestra, indican que, a pesar de tener piernas relativamente largas para su estatura (entre 140 y 165 cm), su constitución era delgada. Además, parece que no estaba bien adaptado para la carrera de resistencia, ya que las pequeñas articulaciones de sus extremidades podrían haber soportado mal las tensiones repetidas y constantes asociadas a esta actividad (Chapman et al., 2025).
- Se ha publicado la revisión del preprint sobre los posibles enterramientos de Homo naledi, reforzando esta hipótesis (Berger et al., 2025).
- A partir de la estructura de los canales semicirculares en el oído interno analizada en una muestra de preneandertales (Sima de los Huesos, SH), neandertales basales (Krapina) y neandertales clásicos de distintos periodos y geografías, se ha podido concluir que los neandertales clásicos poseían una diversidad morfológica notablemente inferior en comparación con los preneandertales y neandertales basales. Esta disminución en la variación fenotípica concuerda con estudios paleogenéticos previos que identificaron un cuello de botella en dicha transición, justo después del máximo térmico de hace 120 ka, al inicio del último ciclo glacial. En cambio, entre los preneandertales de SH y los neandertales basales de Krapina hay similar diversidad morfológica, lo que sugiere continuidad en el linaje neandertal durante el Pleistoceno medio, de forma que en el origen de los neandertales no existió una drástica reducción de su diversidad genética debida a un cuello de botella poblacional (Urciuoli, Martínez, Quam et al., 2025).
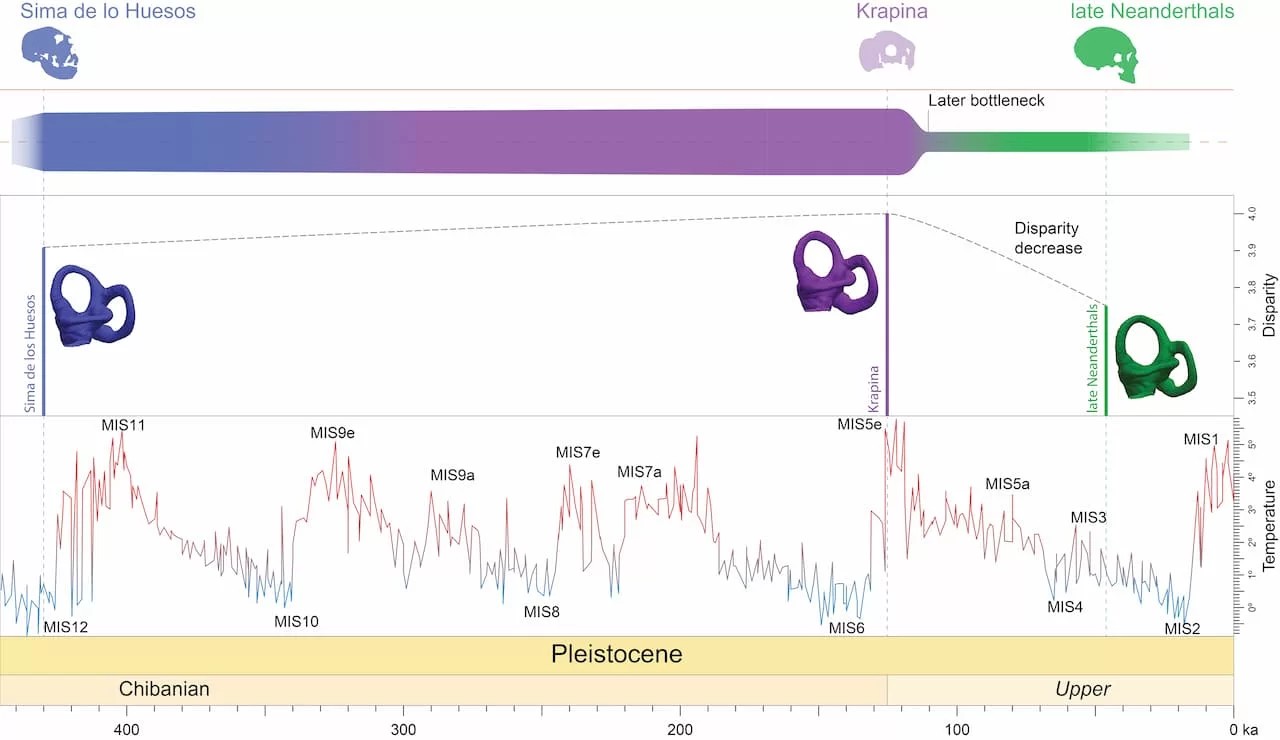
Cambios en la diversidad morfológica de los canales semicirculares a lo largo de la historia evolutiva del linaje neandertal. Crédito: Institut Català de Paleontologia / Miquel Crusafont.
- Un trabajo de Leder y Milks (2025) plantea la hipótesis de que la capacidad tecnológica para el uso de armas de caza a distancia existiría desde hace al menos 300 ka, argumentando que las geometrías de las puntas de lanza encajarían tanto en las lanzas de tipo estoque como en las de tipo jabalina.
- Un estudio genético, apoyado en un nuevo algoritmo computacional llamado Cobraa (Cousins, Scally y Durbin, 2025), revela que Homo sapiens desciende de al menos dos poblaciones ancestrales que se separaron hace 1,5 Ma y se recombinaron hace unos 300 ka, en una proporción aproximada de 80%-20%. La población ancestral mayoritaria sufrió un cuello de botella tras la divergencia, reduciéndose drásticamente en número. Aunque el 20% de la información genética en los humanos actuales proviene de la segunda población, las variantes genéticas heredadas de ella no se distribuyeron al azar en la recombinación: muchos de los genes afectados no están vinculados con regiones funcionalmente importantes, lo que sugiere una selección que eliminaría los elementos menos compatibles, y parece haber habido una fuerte selección que favoreció o eliminó ciertos segmentos, algunos relacionados con la función cerebral y el procesamiento neuronal, lo que plantea incógnitas interesantes respecto a su posible influencia sobre nuestra capacidad cognitiva. Además, existe una fuerte correlación entre las regiones de ascendencia poblacional mayoritaria y la divergencia sapiens-neandertal y sapiens-denisovano, lo que sugiere que la población mayoritaria también era ancestral de esos humanos arcaicos.

Modelo simplificado de la historia demográfica humana según Cousins, Scally y Durbin (2025).
- Se ha datado en unos 150 ka la cultura material del Pleistoceno medio final hallada en bosques tropicales húmedos en el sitio de Beté 1, en el sur de Costa de Marfil (Ben Arous, Blinkhorn, Elliott et al., 2025), mediante la combinación de las técnicas de datación por Luminiscencia ópticamente estimulada (OSL) y Resonancia de paramagnética electrónica (ESR). Es importante la investigación paleoantropológica en las selvas tropicales de África central y occidental, considerando la falta de secuencias arqueológicas y cronológicas completas en la zona.
- Miyagawa y otros (2025) proponen que los estudios genómicos permiten estimar con bastante precisión el momento en que la capacidad lingüística habría surgido en el linaje humano moderno. Para ello, consideran diversos trabajos de investigación genética basados en polimorfismos de un solo nucleótido, que indican que una división poblacional inicial dentro de Homo sapiens a partir del tronco original hace unos 135 ka; y asumen que tanto esta como las divisiones posteriores dieron lugar a poblaciones con plena capacidad lingüística. Así, sugieren que dicho umbral mínimo de 135 ka marca el momento en que el potencial para el lenguaje ya debía estar presente, antes de que ocurriera la primera división poblacional. Si la capacidad lingüística hubiera surgido más tarde, cabría esperar la existencia de algunas poblaciones humanas modernas sin lenguaje o con modos de comunicación fundamentalmente diferentes. Además, se vincula este desarrollo con la aparición generalizada del comportamiento humano moderno, hace unos 100 ka.
- Las interacciones entre grupos sapiens y neandertales en Levante hace 130-80 ka dieron como resultados una homogeneidad en determinadas costumbres sociales y culturales: la tecnología lítica, la mayor dependencia de la caza mayor, los comportamientos de enterramientos intencionales y el uso de ocre. Esta propuesta (Zaidner, Prévost, Shahack-Gross et al., 2025) resulta del estudio de los restos humanos y arqueológicos de la cueva de Tinshemet (Israel), de unos 100 ka, y su comparativa con otras cuevas de la región. En concreto, destacan las prácticas de enterramiento compartidas con Qafzeh y Skhul: los cuerpos enterrados en posición fetal, acostados de lado con las piernas muy flexionadas, los brazos doblados hacia el pecho y la cara, y la cabeza hacia abajo. En Tinshemet se han hallado 7500 fragmentos de ocre de diferentes colores y texturas, algunos transportados desde fuentes distantes, lo que implica un transporte intencional de este material desde decenas de kilómetros para aquel uso simbólico. Recordemos que en esta región vivieron tres grupos humanos distintos: neandertales, humanos modernos y un grupo representado por la mandíbula de Nesher Ramla. Nuevos trabajos seguirán profundizando en determinar hasta qué grado la mencionada homogeneidad cultural provino de interacciones sociales o bien de hibridación entre las especies.

Enterramientos en Tinshemet, Qafzeh y Skhul. Crédito: Zaidner, Prévost, Shahack-Gross et al. (2025)
- Hace unos 90 ka, la población neandertal que usaba la Cueva del Camino, en el valle alto del río Lozoya (Madrid, España) tenía hasta 34 individuos. Los restos faunísticos de la capa 05 indican que era un lugar con un ecosistema muy fértil con mucha biodiversidad, comparable a la de algunos conjuntos faunísticos del Pleistoceno en la Sierra de Atapuerca, como Gran Dolina y Galería, y al Parque Nacional del Serengeti en la actualidad. Entre otras especies, se han identificado gamos, jabalíes, castores, caballos salvajes, uros y rinocerontes esteparios. Esta riqueza natural y la estructura cerrada del valle, rodeado de montañas, permitieron a este grupo de neandertales establecerse y no tener la necesidad de desplazarse a grandes distancias para cazar y recolectar (Molino, Rodríguez-Gómez, Karampaglidis et al., 2025).
- Se han examinado los grabados sobre cinco artefactos de cuevas levantinas: un núcleo centrípeto de Levallois de Manot (68 ka), dos láminas corticales de Amud (55 ka), un núcleo de Qafzeh (100 ka) y una plaqueta grabada de Quneitra (54 ka). Los grabados de los artefactos de Manot, Qafzeh y Quneitra son deliberados, mientras que las marcas en los objetos de Amud son de origen abrasivo. Las incisiones deliberadas muestran formas geométricas similares y tienen patrones alineados con la topografía y la forma de la superficie del artefacto (Goder-Goldberger, Marreiros, Paixão et al., 2025).

Núcleo grabado de Manot. Créditos: foto E. Ostrovsky, esquema M. Smelansky, modelos 3D E. Paixão y L. Schunk.
- La variante I197V de la proteína NOVA1 solo está presente en humanos modernos, no en neandertales ni denisovanos. Esta variante afecta la regulación de genes en el cerebro y puede haber influido en la forma en que procesamos el habla. Tajima, Vargas, Ito y otros (2025) han publicado la edición mediante CRISPR del genoma de ratones, observando múltiples picos de unión en las transcripciones del cerebro de ratón involucradas en la vocalización, es decir, que los ratones con esta variante presentaban diferencias conductuales en los patrones de los sonidos que emitían. Este hallazgo les lleva a sugerir que NOVA1 fue protagonista en la evolución del habla, y que esta sustitución específica para humanos pudo haber sido parte de un evento de selección evolutiva en una población ancestral de Homo sapiens, posiblemente contribuyendo al desarrollo del lenguaje hablado a través de la regulación diferencial del ARN durante el desarrollo de circuitos neuronales.
- Abood y Oota (2025) analizan los distintos trabajos genómicos que describen las dispersiones de los humanos modernos hacia Asia oriental, y las adaptaciones fisiológicas que habían sido necesarias. Cualquiera que fuera la ruta elegida, las migraciones siempre ocurrieron hacia latitudes más altas, lo que requería una adaptación a menor luz, climas más fríos y mecanismos fisiológicos para contrarrestar la escasez de alimentos. La introgresión de denisovanos y neandertales desempeñó un papel importante en las adaptaciones fisiológicas, como la resistencia al frío, el metabolismo y la función inmunitaria. El estudio investigó dos posibles rutas migratorias: una ruta septentrional al norte del Himalaya, y una ruta meridional que atraviesa el subcontinente indio y el sudeste asiático. La meridional habría sido la principal ruta de migración de los antepasados de los asiáticos orientales, incluidos los chinos Han, los japoneses (excluyendo la ascendencia Jomon) y muchas poblaciones del sudeste asiático. El análisis genómico del pueblo Jomon, una antigua población de Japón, indicó que los asiáticos orientales modernos no presentan contribuciones genéticas significativas de la ruta norte, a diferencia de los nativos americanos y los asiáticos del noreste. El grupo del noreste incluye a siberianos, poblaciones mongolas y algunos grupos indígenas de China, Corea y Japón. El estudio también observa que las poblaciones africanas poseen la mayor variación genética, lo que apoya el origen africano de todos los humanos modernos.
- El estudio de los materiales recuperados en Longtan (suroeste de China) en 2019 y 2020, con un total de 3487 artefactos de piedra, ha dado lugar a la identificación de 53 raspadores de Quina y 14 núcleos, los primeros ejemplos de esta industria en el este asiático (Ruan et al., 2025). Este conjunto tiene unos 50-60 ka. ¿Desarrollaron los denisovanos esta tecnología, o pudieron llegar los neandertales europeos hasta esa región?
- Se han hallado restos virales en lecturas de secuencias de datos del genoma de dos neandertales de 50 ka de la cueva Chagyrskaya, Siberia, que incluyen adenovirus, herpesvirus y papilomavirus. En los humanos modernos, estos virus están asociados con resfriados, herpes labial, verrugas genitales y algunos tipos de cáncer, aunque no está claro si los síntomas serían similares en neandertales. Estas infecciones podrían haber inducido respuestas inmunológicas, aunque sería importante el posible debilitamiento de las capacidades físicas en poblaciones reducidas como aquella (Ferreira et al., 2025).
- La gracilidad de la cara humana moderna parece deberse a un cambio en el desarrollo de la actividad celular ósea en la región mediofacial, que comienza a ralentizarse en la infancia y se detiene alrededor de la adolescencia. Esto contribuye a tener una cara pequeña en la edad adulta, en contraste, por ejemplo, con los neandertales y los chimpancés (Schuh et al., 2025).
- La colección de restos neandertales de Grotte du Bison, Arcy-sur-Cure (Francia) está compuesta por al menos entre 9 y 17 individuos, la mayoría niños y adolescentes, que vivieron a comienzos del MIS3, hace unos 57 ka (Henrion et al., 2025).

Colección de dientes de Grotte du Bison. Crédito: Juliette Henrion et al (2025)
- Se ha anunciado el hallazgo de 438 artefactos líticos de industria musterienses con bifaces diversas tipologías en Enclos (meseta de Chartres, Francia) fabricados en sílex local, con datación entre 57,94 y 42,82 ka. En este conjunto se han identificado diversas técnicas de talla, incluyendo Levallois, laminar volumétrica, extracción sobre lascas y tal vez discoide.
- Se ha publicado un completo estudio del ADN sedimentario antiguo (sedaDNA)de la secuencia estratigráfica arqueológica inferior de la cueva de El Mirón (Cantabria, España), que abarca el Musteriense tardío, Gravetiense (31,5 ka), Solutrense (24,5-22 ka) y Magdaleniense inicial (21-20,5 ka). El trabajo (Gelabert, Oberreiter, Straus et al., 2025) ha permitido identificar 28 taxones animales, incluido Homo sapiens. 15 de ellos no habían sido identificados a partir del registro arqueozoológico, como la presencia de hienas en el Magdaleniense. Se ha realizado el análisis filogenético de 70 muestras de ADNmt de fauna, incluido el muestreo más denso del Pleistoceno ibérico de Canis lupus. Finalmente, se han secuenciado tres muestras de ADNmt humano de los niveles solutrenses, que sugieren la continuidad del haplogrupo de ADNmt en Iberia a lo largo del período Solutrense/Último Máximo Glacial.
- Los trabajos en 30 afloramientos estratigráficos cuaternarios en la región de Río Campo o Río Ntem (Guinea Ecuatorial) han producido el descubrimiento de 16 sitios con 418 herramientas de piedra que muestran ocupaciones humanas hace entre 44-20 ka en África central. Estos hallazgos reflejan un conocimiento técnico avanzado y una capacidad extraordinaria de adaptación cultural y organización social para ocupar sistemáticamente las selvas ecuatoriales africanas, uno de los ecosistemas más desafiantes del planeta. Los atributos tecnotipológicos de las herramientas se asemejan a las tradiciones de la MSA y apuntan a una continuidad cultural que se remontaría a unos 250-300 ka (Rosas et al., 2025).
- Un preprint analiza la desaparición de Homo floresiensis y las posibles causas del abandono de Liang Bua hace entre 61-47 ka, apuntando a una aridificación de paisaje y una intensificación de la caza de la fauna local, en un contexto de recursos cada vez más escasos (Gagan et al., 2025).
- La dentición del maxilar de Mugharet El’Aliya (Marruecos), de gran tamaño, junto con la morfología aparentemente arcaica del hueso, llevó a atribuirlo al linaje neandertal (Şenyürek, 1940), si bien revisiones más recientes del fósil habían sugerido afinidades con Homo sapiens. La reevaluación de sus dientes (Röding et al, 2025) confirma su atribución a un H. sapiens juvenil del Ateriense, datado en 35-60 ka, reforzando las evidencias que conectan los especímenes del Ateriense norteafricano con algunos de Asia occidental, especialmente Qafzeh. El canino megadonto y el P4, aunque inusuales para H. sapiens, son típicos del Ateriense. En cualquier caso, dada la posición geográfica del norte de África como posible corredor de dispersión a través de África y Europa, no se descarta que la combinación de rasgos observada puede indicar una posible mezcla con neandertales, aunque también puede responder a otros procesos evolutivos, incluyendo, por ejemplo, la retención de rasgos ancestrales, la adaptación a entornos locales y la deriva genética en poblaciones marginales.
- La comparación de las cajas torácicas del neandertal Shanidar 3 con otro neandertal, Kebara 2, y con 58 Homo sapiens de distribución mundial, indica que el tórax de Shanidar 3 y de Kebara 2 tienen la típica forma de campana descrita para los neandertales, y forma que se separa de la de H. sapiens, siendo más cercanas a los individuos adaptados al frío (López-Rey, García-Martínez y Bastir, 2025). Sin embargo, las similitudes entre Shanidar 3 y Kebara 2, ambos habitando localidades cálidas del Levante durante el Pleistoceno superior, podrían desafiar la idea convencional de un bauplan adaptado al frío en los neandertales.
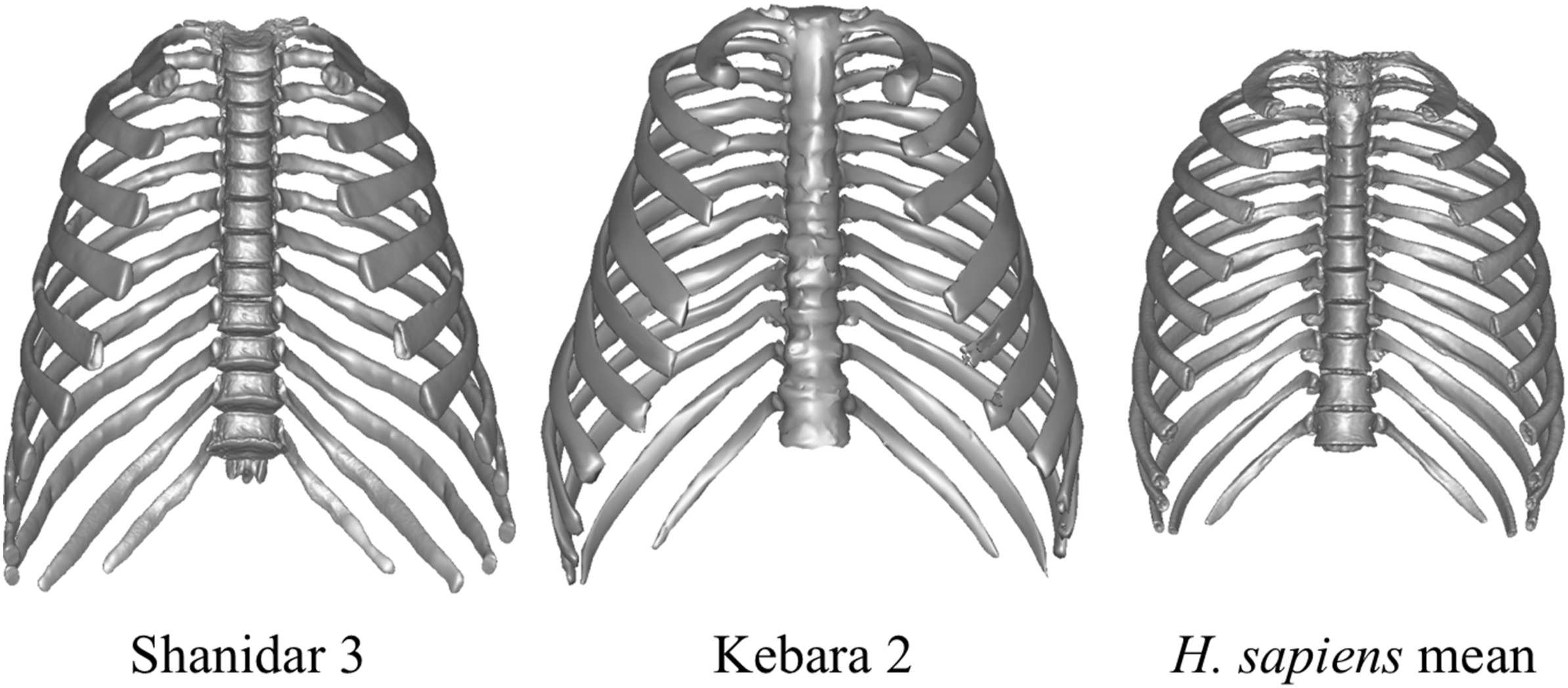
Cajas torácicas de Shanidar 3, Kebara 2 y Homo sapiens. Crédito: López-Rey, García-Martínez y Bastir (2025)
- Se ha estudiado la diversidad genética de los grupos sanguíneos de 22 Homo sapiens y 14 neandertales que vivieron entre 120 ka y 20 ka (Mazières, Condemi, El Nemer et al., 2025). La diversidad de alelos de grupos sanguíneos en los neandertales permaneció inalterada desde hace 120 ka, mientras que los H. sapiens que se expandieron por Eurasia tenían alelos de grupos sanguíneos que actualmente son exclusivos de poblaciones no africanas. Estos cambios rápidos posiblemente ocurrieron hace entre 70 y 45 ka, justo después de su salida de África, y les ayudaría a sobrevivir en diferentes climas y a resistir enfermedades. Además, el sapiens individuo de Ust’Ishim poseía alelos desconocidos que pueden ilustrar la herencia genética perdida de los primeros euroasiáticos. Por último, los neandertales compartían un haplotipo Rh único que sigue presente en algunas personas de Oceanía, a quienes probablemente llegaría a través de los cruces entre H. sapiens y neandertales.
- Se ha logrado determinar con precisión la antigüedad del “Niño de Lapedo”, mediante datación directa de los restos óseos del individuo, resultando entre 27,78 y 28,85 ka (Linscott et al., 2025). Se trata de una datación por radiocarbono con hidroxiprolina, un aminoácido que desempeña un papel crucial en la estructura y función del colágeno de los huesos, y que permite eliminar contaminantes que afectan los resultados de otras técnicas de datación, resultando una técnica mejorada para muestras mal conservadas. Las primeras estimaciones eran de unos 24,5 ka (Duarte et al, 1999). Este esqueleto casi completo (cuya codificación formal es Lagar Velho 1) perteneció a un niño de unos 4 o 5 años de edad al morir, descubierto en 1998 en el centro de Portugal. Estaba teñido de ocre rojo y acompañado de conchas perforadas y huesos de animales, sugiriendo un enterramiento ritual. Rasgos de su morfología hicieron plantear la posibilidad de una hibridación con neandertales: mientras que su cráneo tiene rasgos sapiens, sus extremidades inferiores eran cortas y robustas, más similares a las de un neandertal. Sin embargo, su datación reciente reduce esta posibilidad (Tattersall y Schwartz, 1999), si pensamos que los últimos neandertales desaparecieron de Iberia hace unos 40 ka, con permiso de las intrigantes dataciones de 33-24 ka para los últimos niveles musterienses de Gibraltar (Finlayson et al., 2008) que serían contemporáneas al Niño de Lapedo.

Esqueleto del Niño de Lapedo. Crédito: João Zilhão y Cidália Duarte
- Se ha anunciado en prensa el interesante enterramiento de un niño de hace al menos 29 ka en Tham Din (Cueva de Tierra), Tailandia. En esta cueva, se conocían pinturas en ocre rojo en las paredes y el techo de tres cámaras que representaban personas, algunas usando arcos y flechas para cazar animales como ciervos y monos. En el yacimiento también se han hallado huesos de animales, conchas, semillas y artefactos líticos. Bajo estos, apareció un niño humano moderno de entre 6 y 8 años, tendido boca arriba, con los brazos y las piernas pegados al cuerpo, lo que indica que probablemente fue enterrado envuelto o atado. También existen restos de una hoguera junto al cuerpo. Esperaremos su publicación.
- Atendiendo al desgaste dental, es probable que hace unos 25-29 ka los pueblos de la cultura pavloviense, una variante regional del Gravetiense en Europa central, usaran labrets o piercing labiales (Willman, 2025).
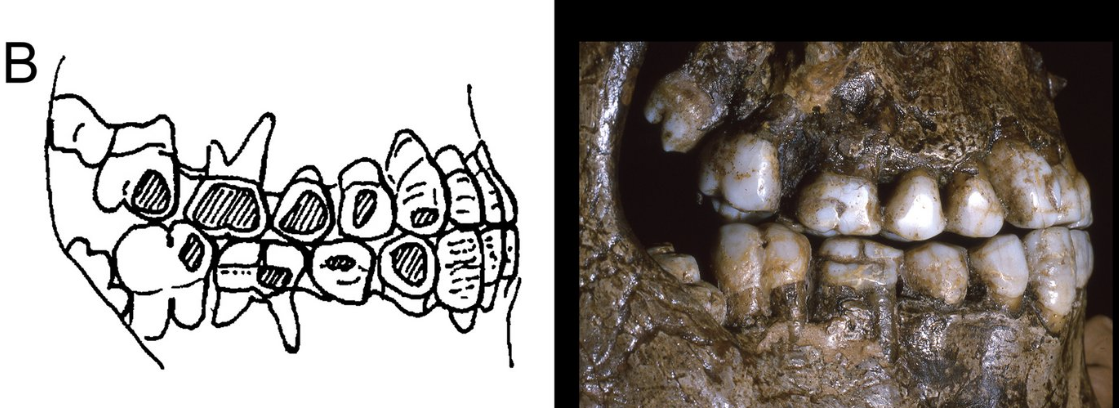
Uso de labrets en la cultura pavloviense. Crédito: J. C. Willman.
- Bennett y otros (2025) interpretan unos rastros en el Parque Nacional White Sands de Nuevo México (EE. UU.) como dejados por uno o varios postes de madera asociados a las pisadas humanas, que corresponderían a un dispositivo simple de transporte denominado travois, una especie de carretilla sin ruedas.

Área A de White Sands con rastros lineales y huellas humanas. Crédito: Bennett et al., 2025
Se identifican incluso diferentes estilos, unos que dejaban dos líneas (dos postes en forma de X) y otros que dejaban una sola línea (compuestos por dos piezas de madera formando un triángulo).

Reconstrucción de los dos tipos de travois posiblemente utilizados hace 22 ka en White Sands
- Utilizando micromorfología, fluorescencia de rayos µ-X y microscopía electrónica de barrido, se han reconstruido múltiples fases de visitas humanas durante el Gravetiense a la cueva de Fuente del Salín (Cantabria, España), mostrando un uso intensivo del fuego, como lo indican los abundantes huesos y conchas marinas calentados, carbones, derivados de grasas y restos in situ de posibles hogueras abiertas apiladas, así como lechos de hierba quemada. La quema intensiva, la reutilización sistemática de elementos de combustión y los múltiples propósitos de los fuegos en Fuente del Salín son comparables con los de los yacimientos del Gravetiense de Europa central y oriental, lo que indica que estos comportamientos de uso del fuego probablemente no reflejen una adaptación regional a los entornos periglaciares, sino un rasgo cultural de la tradición del Gravetiense en toda Europa (Alzate-Casallas, Sánchez-Carro, Barbieri y González-Morales, 2025).
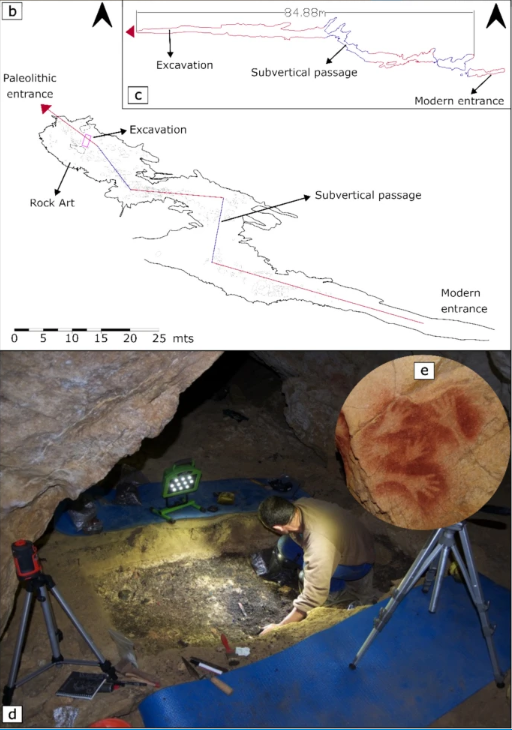
Fuente del Salín: b) Excavación, arte rupestre, entradas moderna y paleolítica. c) Sección longitudinal y topografía. d) Excavación. e) Huellas de manos en rojo en negativo en el interior de la cueva asociadas a los cazadores-recolectores del Gravetiense. Crédito: Alzate-Casallas, G., Sánchez-Carro, MA, Barbieri, A. et al. (2025)
- El estudio del aprovisionamiento de sílex en los periodos gravetiense, proto-solutrense y solutrense en Vale Boi, Portugal (Belmiro, Terradas, Dominguez-Bella y Cascalheira, 2025) revela que la mayor parte es de origen local, dentro de un radio de 20 km y, en menor medida, de fuentes no locales situadas a más de 200 km en el centro de Portugal y el sur de España.
- Se han documentado acciones de canibalismo hace 18 ka en la cueva de Maszycka, Polonia (Marginedas, Saladié, Połtowicz-Bobak et al., 2025). El 68% de los 63 fragmentos óseos humanos (de 6 adultos y 4 niños) presentan cortes y fracturas intencionadas asociadas al consumo humano, como la extracción de músculos, cerebro y médula ósea. Una posible interpretación de este hallazgo es la selección de cráneos como práctica funeraria, también observada en otros conjuntos magdalenienses en Europa continental, tal vez en el marco de enfrentamientos intergrupales, causados por la competencia por los recursos en la expansión demográfica tras el Último Máximo Glacial. La presencia humana documentada en Maszycka parece corresponder a ocupaciones efímeras. El perfil de edad de los restos se asemeja al de una unidad familiar nuclear completa y, a diferencia de lo observado en Gough’s Cave (Reino Unido) y Brillenhöhle (Alemania), donde los cráneos se manipularon para fabricar “cráneos copa” rituales, en Maszycka no se han encontrado signos de un tratamiento ritual especial, lo que sugiere que el grupo de Maszycka podría haber sido atacado, sometido y canibalizado. Los conjuntos magdalenienses con evidencia de canibalismo parecen sugerir que esta práctica no era especialmente extraordinaria, aunque tampoco un fenómeno común u homogéneo durante este periodo, es decir, no sería una parte habitual de su vida cotidiana o doméstica. Por ahora, se han identificado 25 casos de canibalismo en Europa, abarcando el último millón de años, siendo al menos cinco del Magdaleniense.
- Los grabados del abrigo paleolítico Ségognole 3 (Noisy-sur-École, Francia) podrían interpretarse como un mapa tridimensional de la zona realizado hace unos 13 ka (Thiry y Milnes, 2025). Allí ya se habían documentado los grabados de dos caballos a ambos lados de una vulva, a la cual llegaba agua canalizada gracias a la modificación de las fracturas de las paredes. A dos metros de estas representaciones, el suelo de arenisca también fue modificado para representar las elevaciones del paisaje más cercano y sus flujos de agua natural, en el valle del río École. Hasta ahora, el mapa tridimensional más antiguo conocido era una gran losa de roca grabada de hace 3 ka, en la Edad del Bronce.
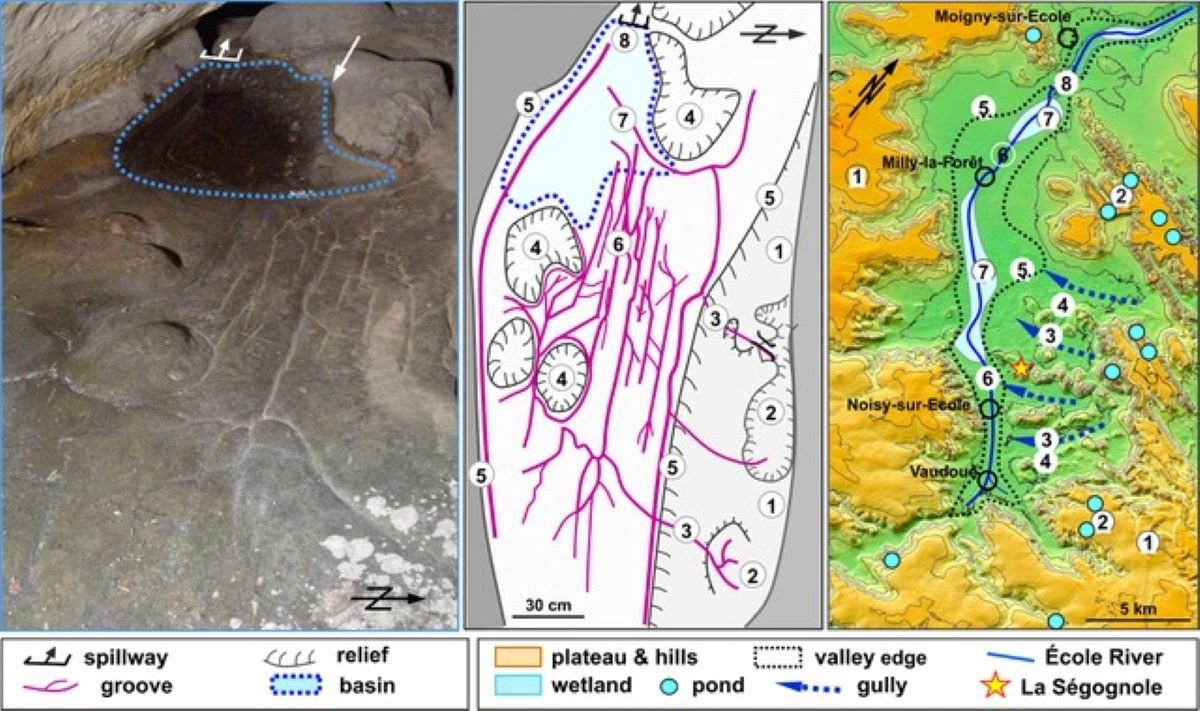
Mapa 3D del abrigo de Ségognole 3. Crédito: Médard Thiry
- Se ha anunciado en prensa el hallazgo de un asentamiento prehistórico de hace 11 ka cerca de la comunidad indígena Sturgeon Lake, en la provincia canadiense de Saskatchewan. Se trataba de una sociedad permanente, de la que se han encontrado herramientas líticas, materias primas, huesos de megafauna procesada y restos de hogueras. Este descubrimiento indica la existencia de sociedades organizadas en la región mucho antes de lo esperado, donde se pensaba que los grupos en esta antigüedad eran nómadas.
- La evidencia más antigua conocida de la mezcla de ingredientes para la elaboración de veneno destinado a puntas de flecha tiene 7 ka y se ha encontrado en la cueva Kruger (Sudáfrica). En un fémur de bóvido se hallaron tres puntas de flecha de hueso y restos de digitoxina y estrofantidina, glucósidos cardíacos relacionados con venenos de caza (Bradfield, Dubery, Steenkamp, 2025). También se identificaron restos de ácido ricinoleico, que pudo una tercera toxina. En una herramienta de madera de 24 ka hallada en la cueva Border (Sudáfrica) se observó también este compuesto, que pudo ser el componente de un veneno, aunque esta interpretación se contestó, dado que pudo también tener otros usos.
- A partir de 350 genomas antiguos de Eurasia que vivieron entre hace 45 y 1,7 ka, un preprint (Perretti et al., 2025) aporta nueva información sobre cómo evolucionó el color de la piel, los ojos y el cabello. El cambio hacia pigmentaciones más claras resultó ser prácticamente lineal en tiempo y espacio, y más lento de lo esperado. Las primeras variaciones genéticas asociadas con la piel clara aparecieron hace unos 14 ka, pero su presencia era minoritaria. La mayoría de individuos tendrían colores de piel oscuro o intermedio hasta hace unos 3 ka. No fue un fenómeno homogéneo, sino que se produjo a diferentes ritmos en distintas regiones, con una gama de tonalidades que empezó a diversificarse significativamente en la Edad del Bronce y alcanzó su punto álgido en la Edad del Hierro. La explicación tradicional de la evolución de la piel clara se basa en la necesidad de producir vitamina D en latitudes con menor radiación solar. Sin embargo, si las poblaciones antiguas consumían alimentos ricos en esta vitamina, como pescado y carne, la presión evolutiva para desarrollar piel más clara podría haber sido menor. La agricultura y la reducción del consumo de carne y pescado harían de la piel más clara una ventaja adaptativa. Por otra parte, los ojos claros aparecieron antes de que la piel clara se volviera común. En el Mesolítico existió un pico de pigmentación clara en los ojos y un cambio acelerado durante la expansión de los agricultores neolíticos por Eurasia occidental. Los ojos azules podrían haber surgido por selección sexual o simple deriva genética, sin un beneficio adaptativo claro.
- El estudio del genoma de 57 individuos de la tribu Durotriges del final de la Edad del Hierro en el sur de Gran Bretaña, indica que eran sociedades matrilocales en que las mujeres permanecían de por vida en la comunidad y los hombres se movían entre poblados (Cassidy, Russell, Smith et al., 2025).
- Se ha anunciado el conjunto de herramientas de producción de fuego procedente de Caoyangang, en la provincia de Jiangsu (este de China), de una antigüedad estimada de 7000 años. Se trata de un palo de perforación de 60 cm de largo y una tabla de 30 cm de largo, que contiene 10 hendiduras circulares con claros signos de quemado y una ranura circular en un extremo, probablemente utilizada para atar una cuerda, lo que sugiere que fue diseñado para transportarlo o colgarlo.

Conjunto de herramientas para producir fuego de Caoyangang. Crédito: Xinhua News
- Se ha confirmado la antigüedad de las pinturas rupestres con representaciones navales del Abrigo de Laja Alta (Cádiz), uno de los escasos testimonios de navegación prehistórica de la península ibérica (Morgado-Rodríguez et al.,2018 y 2025). Se han obtenido dataciones absolutas del IV-III milenio a.C., durante el Neolítico reciente y Calcolítico del sur de Iberia.

Representaciones navales prehistóricas del abrigo de Laja Alta (Jimena de la Frontera, Cádiz). Crédito: Morgado-Rodríguez et al., 2025.
- Una investigación de Ma, Lu, Stoneking y Xu (2025) ha encontrado una posible explicación genética a la persistencia de la lactasa, que motiva que los humanos de ascendencia europea mayoritariamente pueden digerir fácilmente la lactosa, frente a los humanos de ascendencia asiática oriental. El 25 % de las muestras de Asia oriental estudiadas presentaban versiones únicas del gen de la lactasa, que no se encuentran en los genes de personas africanas o europeas. Además, estas variantes genéticas en los asiáticos orientales probablemente provenían de los neandertales, quienes podrían haber desarrollado esta adaptación genética como consecuencia de algún tipo de protección contra las infecciones.

Eres fabuloso! Dios te bendiga por trabajar para bien de los demás ??????
Enviado desde Outlook para Androidhttps://aka.ms/AAb9ysg ________________________________
Me gustaLe gusta a 1 persona
Gracias. Muchísimas gracias por todo lo que compartes. Más gente como tú.
Belén
Me gustaLe gusta a 1 persona
Una impressionante mole di notizie, che deve aver richiesto non poco tempo e lavoro per essere sintetizzata e proposta. Per un semplice appassionato settantenne non addetto ai lavori è una delle pochissime possibilità di essere aggiornati, una fonte di fatto unica nel panorama mondiale, davvero grazie dr. Saez!
Mi scusi se Le scrivo in italiano ma non ho altre possibilità.
Lo studio genetico di Cousins, Scally e Durbin è di grande impatto. Ho presente uno schema di John Hawks, ormai di 5 anni fa ( https://x.com/johnhawks/status/1230874819366772739 ), con differenze sostanziali rispetto a questo recente lavoro. Ho letto un paio di volte di accenni ad una ibridazione dei soli africani (a differenza dello studio attuale) con una popolazione più arcaica, aspetto di cui si fa cenno in questa pubblicazione.
L’idea di una lunghissima separazione (1 milione e 200.000 anni!) in Africa di due popolazioni umane non è facile da immaginare / visualizzare, forse un gruppo si era specializzato in ambiente forestale? Le date rendono spontaneo pensare a Homo naledi, tuttavia con differenze tali (dimensioni del cervello, corporatura, adattamento all’arrampicata, ecc.) rispetto ai quasi – sapiens di 300.000 anni fa da porre molti dubbi su questa eventuale ibridazione.
Si tratta anche di una ibridazione di grande rilievo (80 – 20%), non di un fenomeno marginale.
A un milione e 500.000 anni fa risalgono, tra gli altri, il Turkana Boy e KNM-ER 3883. È un periodo più freddo ed arido rispetto al passato, che potrebbe aver favorito la separazione tra le popolazioni settentrionali e quelle meridionali dell’Africa (e il possibile incontro con le popolazioni eurasiatiche, sospinte verso sud dai ghiacci). I periodi aridi in Africa orientale potevano rappresentare una barriera tale da mantenere relativamente separate (e per tanto tempo) una popolazione africana settentrionale da una meridionale (magari con fasi di incontro e parziale ibridazione)? Tante domande, sono curioso di vedere quale accoglienza avrà questo studio.
Angelo Mammana
Me gustaLe gusta a 1 persona
Gracias como siempre Roberto, un autentico placer leer tus resúmenes y hojear artículos. Un cordial saludo
Me gustaLe gusta a 1 persona