El origen de la tecnología lítica en la evolución humana despierta un gran interés. Los hallazgos, cada vez más antiguos y complejos, indican que su fabricación no se restringió a nuestro género Homo. Algunos podrían asociarse con ciertos parántropos y australopitecos que coexistieron con los primeros humanos. Sus manos presentan adaptaciones que los acercan a la destreza manual humana mucho más que a la de otros primates. Microcharla impartida para el Club de Ciencia Boadilla el 14 de noviembre de 2025.
Nota: Este artículo se basa en dos preprints pendientes de revisión por pares. Se actualizará con las versiones que sean publicadas.
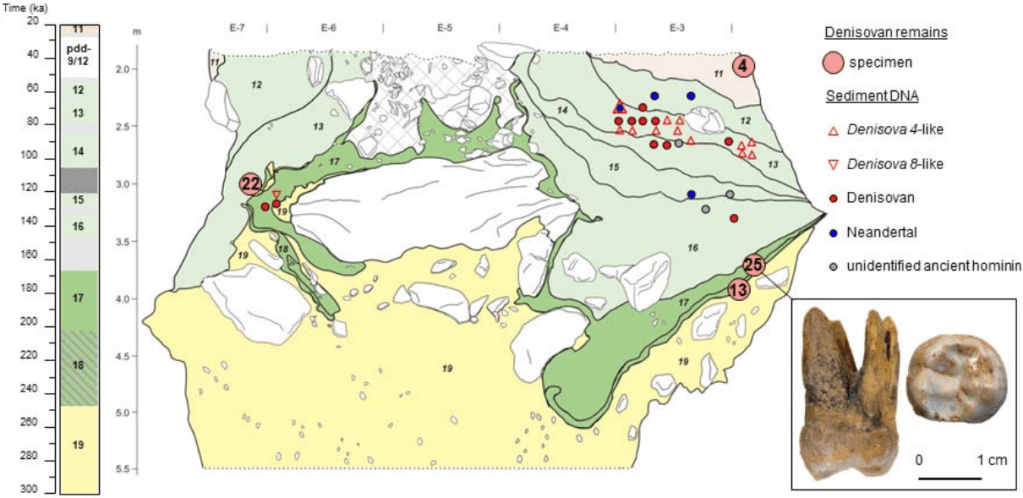
Un trabajo de Peyrégne y colaboradores presenta el genoma de alta cobertura obtenido a partir del esmalte del molar Denisova 25, procedente de la cueva de Denisova (Altái, Siberia). El diente perteneció a un individuo masculino adulto de entre 20 y 35 años, que vivió hace unos 200 ka (miles de años) según las dataciones por luminiscencia del sedimento (OSL). El genoma, extraordinariamente conservado, aporta obtener perspectivas muy interesantes sobre la diversidad genética de las poblaciones denisovanas y su interacción con otros linajes humanos.
Perfil estratigráfico de la cámara sur de la cueva Denisova. Crédito: Peyrégne et al. (2025).
Los análisis de homocigosidad (Runs of Homozygosity, ROH) indican que el grupo al que pertenecía D25 era muy reducido, de unos 50–60 individuos, comparable al de poblaciones paleolíticas extremadamente aisladas. Su población habría divergido de la que dio origen a Denisova 3 hace en torno a 215–230 ka.
Las poblaciones de humanos modernos en el este y sudeste asiático se diversificaron hace menos de 40 ka (miles de años) y presentan ascendencias basales que, en gran medida, fueron reemplazadas por un único linaje, que se diversificó en las ascendencias de la mayoría de los asiáticos actuales. Sin embargo, la diversidad genética observada en muchas poblaciones del suroeste de China no había sido caracterizada en detalle hasta ahora.
Wang y colaboradores han abordado el estudio sobre la provincia de Yunnan, en la intersección de la meseta tibetana, el sudeste asiático y el sur de China, que alberga la mayor diversidad étnica y lingüística de la China actual. Para explicarla, los autores han analizado los datos genómicos de 127 humanos de Yunnan fechados entre hace 7,1 y 1,4 ka (miles de años), obteniendo los siguientes resultados:
1) Un individuo de 7,1 ka del yacimiento de Xingyi, en el centro de Yunnan, revela una ascendencia basal no documentada previamente, profundamente divergente de las ascendencias del este asiático y que persistió a nivel genético en el sur de Asia oriental desde hace al menos 40 ka hasta mediados del Holoceno.
La crianza humana no solo es una cuestión de cuidado, sino también de energía. A lo largo de la evolución, los cambios en cómo crían los humanos y quién colabora en esa tarea han tenido profundas consecuencias biológicas y sociales.
El coste energético de tener descendencia se reduce significativamente cuando los patrones reproductivos humanos adoptaron períodos de lactancia más cortos y menores intervalos entre nacimientos. ¿Comenzó este patrón en los primeros Homo? Aiello y Key (2002) concluyen que, para hacer posible el acortamiento de la lactancia y del intervalo intergenésico (entre nacimientos), y cubrir el requerimiento energético de un cuerpo mayor respecto a los australopitecos, en los primeros humanos debió producirse un cambio fundamental en el comportamiento de subsistencia, que involucrara una dieta de mayor calidad y la cooperación intergeneracional en la adquisición de alimentos.
Las autoras centraron su análisis en los requerimientos energéticos de la gestación y la lactancia, así como en sus implicaciones sobre el tamaño corporal y la disminución del dimorfismo sexual en los australopitecos, Homo erectus y Homo sapiens, examinando patrones de actividad, masa corporal e historias de vida de primates actuales. De forma similar a lo que ocurre en distintos estudios paleoantropológicos, se suele asumir que los australopitecos tenían un patrón reproductivo más próximo al de los chimpancés actuales, mientras que el de H. erectus sería más próximo al de los humanos modernos.
Este escenario evolutivo habría sido un importante motor en el origen de la organización social, basada en la división económica del trabajo y la cooperación de otros individuos, como subadultos, mujeres de mayor edad (lo que lleva a la famosa hipótesis de la abuela, que veremos más abajo) y otros miembros del grupo.
Aiello (2025) ha actualizado recientemente este análisis apoyándose en los últimos desarrollos en investigación del coste energético, y concluye que las interpretaciones originales siguen siendo válidas, aunque los parámetros de modelado se han ajustado conforme a estudios más recientes sobre el aumento del tamaño corporal y cerebral y la evolución del comportamiento de subsistencia cooperativa.
Aunque existe una gran variación en el tamaño corporal de las primeras especies de Homo, la tendencia general es el incremento, y H. erectus tiene un tamaño promedio mayor que los australopitecos y los primeros representantes del género Homo. No está claro por qué se produjo este incremento en el tamaño corporal de los homininos, lo que resultó en mayores costes diarios de energía, pero se han propuesto diversos factores influyentes, como la termorregulación y el equilibrio hídrico, la eficiencia locomotora, la longevidad y la historia de vida, el crecimiento cerebral y la capacidad de madres más grandes de suministrar más energía a descendientes con cerebros mayores.
El yacimiento de Prado Vargas, situado en el complejo kárstico de Ojo Guareña (Burgos, norte de la península ibérica), aporta información clave sobre las últimas poblaciones neandertales antes de la llegada de Homo sapiens a la región. Un estudio de carácter interdisciplinar (Navazo et al., 2021) reconstruye la vida cotidiana y las tradiciones culturales de estos grupos combinando análisis de estratigrafía, cronología, polen, fauna, industria lítica y restos humanos.
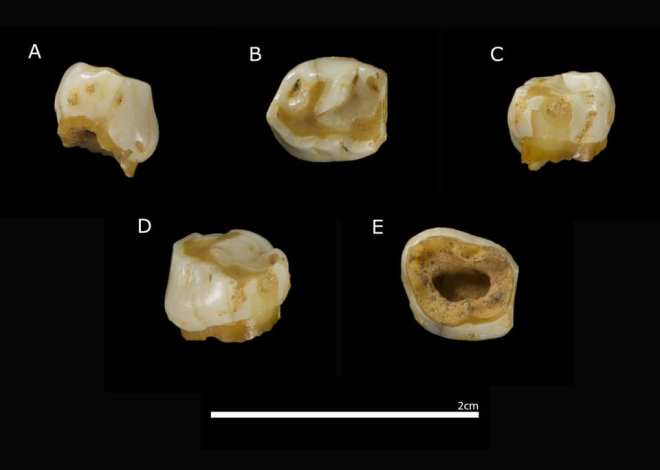
El nivel arqueológico más relevante, N4, se ha datado mediante OSL y radiocarbono entre 54,7 y 39,8 ka (miles de años), por tanto en el MIS 3, un periodo de intensos cambios climáticos. Este nivel se interpreta como un palimpsesto producto de ocupaciones recurrentes, probablemente estacionales (verano-otoño), cuando la cueva permanecía seca. Entre los hallazgos destaca un diente deciduo de un niño neandertal, evidencia directa de la presencia de individuos jóvenes en el grupo.
El diente de Prado Vargas: ¿de una niña o de un niño?
El mencionado diente es un deciduo inferior izquierdo (PV-1360) de un individuo de unos 9-10 años, que lo perdió de manera natural antes de la muerte. Inicialmente se le apodó Vera, que es el nombre de la nieta de Beni, dueño del prado donde se encuentra el yacimiento.
Su conservación ha permitido estudiar las proteínas del esmalte. El esmalte dental funciona como un sistema cerrado, protegiendo las proteínas originales durante decenas de milenios. Entre ellas destaca la amelogenina, codificada por dos genes parálogos: AMELX en el cromosoma X y AMELY en el cromosoma Y. Gracias a la espectrometría de masas aplicada a un pequeño fragmento de esmalte (unos 30 mg), fue posible detectar péptidos exclusivos de AMELY, lo que certifica que el individuo poseía un cariotipo XY. Es decir, cromosómicamente era un individuo masculino.
Este trabajo (Carrillo-Martín, 2025), presentado en el congreso de la ESEB (Evolutionary Society for Evolutionary Biology), constituye un nuevo ejemplo significativo de cómo la paleoproteómica puede suplir la ausencia de ADN en fósiles antiguos, aportando información biológica esencial, como la determinación del sexo o las relaciones filogenéticas.
Ahora bien, ¿cómo rebautizar al joven neandertal…? Yo propondría Vero, a partir del latín verus («verdadero»), para celebrar la determinación de su sexo cromosómico y, en breve, su certificación molecular como neandertal, gracias de nuevo a su proteoma.
Diente neandertal PV-1360. Crédito: Equipo de investigación de Prado Vargas.