

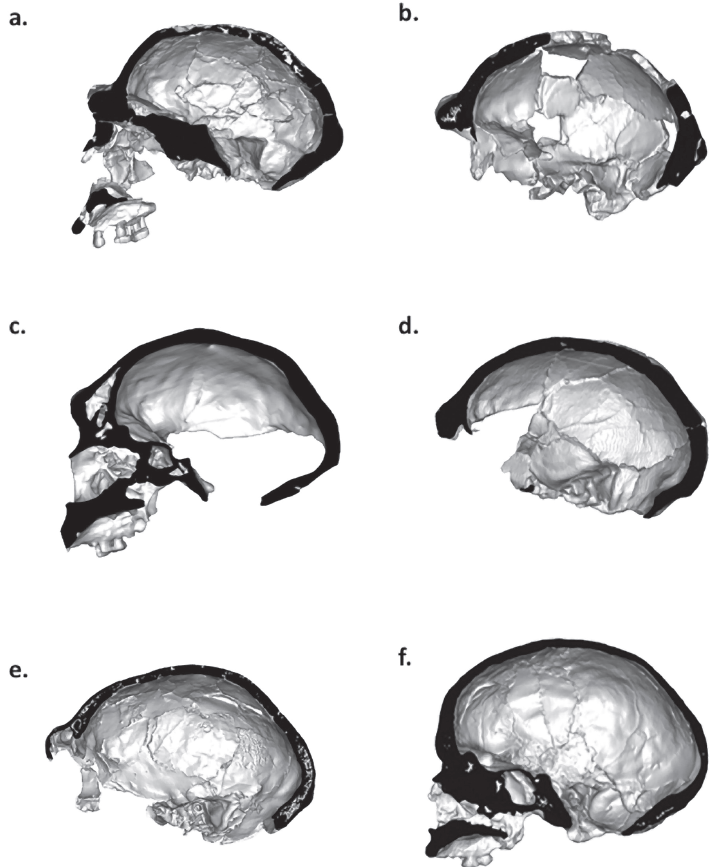
Una de las características más llamativas de Homo sapiens es nuestra frente alta y vertical. La combinación de una serie de rasgos nos distingue frente a otras especies humanas, tales como el cráneo globular, la mencionada frente, el arco superciliar reducido o ausente, la cara retraída, la fosa canina y el mentón pronunciado.
El hueso frontal refleja patrones evolutivos de las poblaciones del Pleistoceno, de forma que H. erectus y H. neanderthalensis tienen frentes inclinadas y toros supraorbitales prominentes, mientras que H. sapiens presenta una frente más vertical y una reducción significativa del torus. Curiosamente, las variaciones suelen ser más evidentes en los laterales del hueso frontal que en su región media.
La evolución de nuestro frontal vertical se ha relacionado con distintos factores, como una combinación de cambios en la base craneal, el crecimiento cerebral y el desarrollo posnatal en nuestra especie.
Cambios en el esfenoides y en la flexión de la base craneal
El esfenoides es el hueso central de la base del cráneo, a partir del cual crece la cara hacia adelante. El acortamiento del esfenoides en nuestra ontogenia, en comparación con el de otros homininos, fue visto desde hace tres décadas como causa de la retracción facial y de una configuración craneal más globular, con un aumento del ángulo entre la base craneal y la parte superior del cráneo. Sin embargo, aunque el acortamiento del esfenoides contribuye al cambio, es insuficiente por sí solo para explicar la totalidad de la transformación hacia una frente alta.
La flexión de la base craneal consiste en el proceso de curvatura que se produce particularmente en la interacción entre el esfenoides y el occipital. La sincondrosis esfeno-occipital es un punto de crecimiento cartilaginoso entre estos huesos, eje sobre el que descansa la biomecánica del cráneo, y juega un papel clave en esta reorganización. En H. sapiens, la prolongación del crecimiento de esta estructura durante la ontogenia permite una mayor adaptación a las demandas del crecimiento cerebral.
Una mayor flexión reduce la longitud de la base anterior, eleva la frente y facilita una reorganización más uniforme del volumen craneal, para alojar un cerebro más globular. El aumento relativo del volumen de los lóbulos temporales también podría estar relacionado con esta flexión, y tener un posible vínculo con funciones cognitivas y lingüísticas asociadas a estas regiones.
Desarrollo posnatal
Se ha observado en distintas especies una correlación entre las vías que sigue el desarrollo posnatal y la altura del neurocráneo. Comparado con sus antepasados, el género Homo experimentó una trayectoria única de desarrollo en la que retiene características juveniles en la etapa adulta, como proporciones neurocraneales elevadas. Especies como H. naledi y H. longi muestran un crecimiento negativo del neurocráneo respecto a la cara, un rasgo arcaico. Este punto de partida ontogenético cambia en H. sapiens. Es evidente que los neandertales y los humanos modernos alcanzan tamaños cerebrales adultos comparables a través de distintas trayectorias de desarrollo, en las evoluciones paralelas de estos linajes «primos».
La expansión del cerebro durante los primeros años contribuye a reorganizar la estructura craneal y al desplazamiento vertical de la frente. Por ejemplo, las fontanelas son espacios abiertos entre los huesos del cráneo que permanecen sin osificar durante los primeros años de vida. En H. sapiens, las fontanelas se mantienen abiertas por más tiempo en comparación con otras especies humanas. Este fenómeno facilita el crecimiento cerebral postnatal y permite un remodelado craneal que favorece la formación de una frente alta.
Reorganización cortical y crecimiento cerebral
El tamaño del cerebro de H. sapiens también importa. Con un volumen cerebral promedio de unos 1350 cc, nuestra especie supera en capacidad a la mayoría de los homininos arcaicos, salvo a H. neanderthalensis y a H. juluensis (denominación que viene a significar «cabeza enorme»). El cráneo juvenil de H. sapiens habría sido un motor de este mayor volumen cerebral relativo y de la frente más vertical, sin requerir un aumento absoluto de la masa encefálica.
En particular, la expansión de los lóbulos frontales genera una mayor demanda de espacio en la región anterior del cráneo. El lóbulo frontal no solo ha aumentado en volumen, sino que también ha sufrido una reorganización funcional, que incluye la elevación de la frente. Por ejemplo, la reconstrucción digital de los cráneos de Buia y Bodo muestra que la forma del frontal no depende exclusivamente del tamaño cerebral, sino de restricciones mecánicas y topográficas del cráneo. Esto afecta al desarrollo de áreas asociadas con la cognición avanzada en la corteza prefrontal.
Factores biomecánicos y tensión funcional
Finalmente, se ha propuesto que la disminución de las tensiones mecánicas en el cráneo también ha podido influir en la morfología de la frente. Las fuerzas generadas durante la masticación crean una presión que puede estar relacionado con el prognatismo facial marcado y la prominencia del torus supraorbital. En H. sapiens, la reducción del prognatismo podría haber reducido parte de estas tensiones, favoreciendo una frente más elevada. Sin embargo, también se ha relacionado el desarrollo del torus supraorbital con cambios en la relación espacial entre el neurocráneo y la cara, más que a tensiones funcionales masticatorias o cargas musculares.
Sigue leyendo
")


