Al mismo tiempo que los Homo erectus comienzan a desarrollarse en África, con los ejemplares más antiguos conocidos datados en 2 Ma (millones de años), otros homininos realizaban marcas de corte en Europa hace 1,95 Ma, según un reciente estudio (Curran, S.C., Drăgușin, V., Pobiner, B. et al., 2025). Este hallazgo se localiza en el yacimiento de Grăunceanu (valle del río Olteț, sur de Rumanía) y representa una antigüedad superior a la de los fósiles de Dmanisi y Orozmani (Georgia), fechados entre 1,85 y 1,77 Ma.
Los ocho especímenes con marcas de corte identificadas con alta confianza incluyen cuatro tibias, una mandíbula, un húmero y dos fragmentos de huesos largos. Todos los restos pertenecen a artiodáctilos, excepto una tibia pequeña de un carnívoro. La mayoría de estos especímenes presentan dos o más marcas lineales atribuibles a cortes. Además, se han identificado otros trece especímenes con posibles marcas de corte, aunque con menor grado de certeza.

Especímenes con marcas de corte de hace 1,95 Ma hallados en Grăunceanu. Crédito: Curran, S.C., Drăgușin, V., Pobiner, B. et al. (2025)
Los siguientes restos humanos más antiguos fuera de África incluyen una vértebra lumbar inferior muy robusta de 1,5 Ma hallada en el sitio israelí de ‘Ubeidiya; un diente deciduo de Barranco León (Orce) y una cara parcial de Sima del Elefante (Atapuerca), ambos en España y fechados en 1,4 Ma; así como la calota craneal de Kocabaş (1,1-1,3 Ma) en Turquía.
Por otro lado, en Asia, la presencia humana más antigua conocida es todavía mayor: herramientas líticas en Shangchen, al norte de China, que datan de 2,1 Ma. Por tanto, es posible que grupos de homininos que comenzaban a desarrollar rasgos físicos y conductuales asociados con la modernidad (o, al menos, así lo etiquetamos hoy) se expandieran rápidamente fuera de África, tanto hacia el oeste (Europa) como hacia el este (Asia), entre hace 2,1 y 2,5 Ma.
En conjunto, la evidencia apunta a una presencia generalizada, aunque posiblemente intermitente, de homininos en Eurasia antes de los 2 Ma. El mapa siguiente destaca en azul los yacimientos conocidos con evidencia de presencia humana anterior a esta fecha.

Sites shown in blue text are suggested to be > 2 Ma. Credit: Curran, S.C., Drăgușin, V., Pobiner, B. et al. (2025)
El mismo estudio ofrece una destacada recopilación de los yacimientos con presencia humana del Pleistoceno inicial en Eurasia y el norte de África anteriores a 1 Ma. En cada caso, se detalla la presencia de restos humanos, materiales líticos y evidencias de modificaciones realizadas por homininos, como marcas asociadas al descuartizamiento.
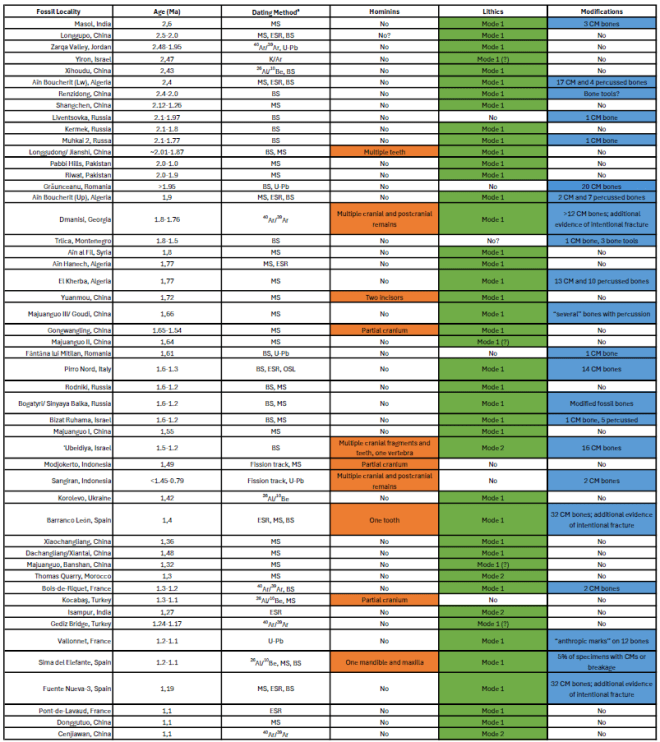
Reported hominin localities from the Early Pleistocene of Eurasia and northern Africa prior to 1 Ma. Credit: Curran, S.C., Drăgușin, V., Pobiner, B. et al. (2025)
Más información:
- Una cara humana de 1,4 millones de años en Europa occidental | Nutcracker Man
- The first Europeans: evidences in Southern Spain | Nutcracker Man
- The first Europeans: summary of key sites and evidences in Western Turkey | Nutcracker Man
- Sobre la expansión humana por Asia, mucho más antigua de lo que pensábamos | Nutcracker Man