The #FossilFriday tradition seems to be losing steam, and even hashtags themselves may no longer be as popular as they once were. This trend is also reflected in the declining number of #FossilFriday posts featuring hominin fossils. Still, as long as there continue to be outstanding contributions like those we’ve seen this year, this Top 10 list will live on!
But what exactly is ‘FossilFriday’? Every Friday, social media users share captivating fossil images under the hashtag #FossilFriday. These posts often showcase iconic specimens, museum collections, scientific papers, blog articles, quirky fossils, and much more. I especially enjoy joining in by posting about hominin fossils. So, without further ado, here’s the Top 10 of 2025. Keep posting!
10. The hypnotizing temporomandibular joint of KNM-ER 23000. By Marta Mirazón Lahr @MartaMLahr

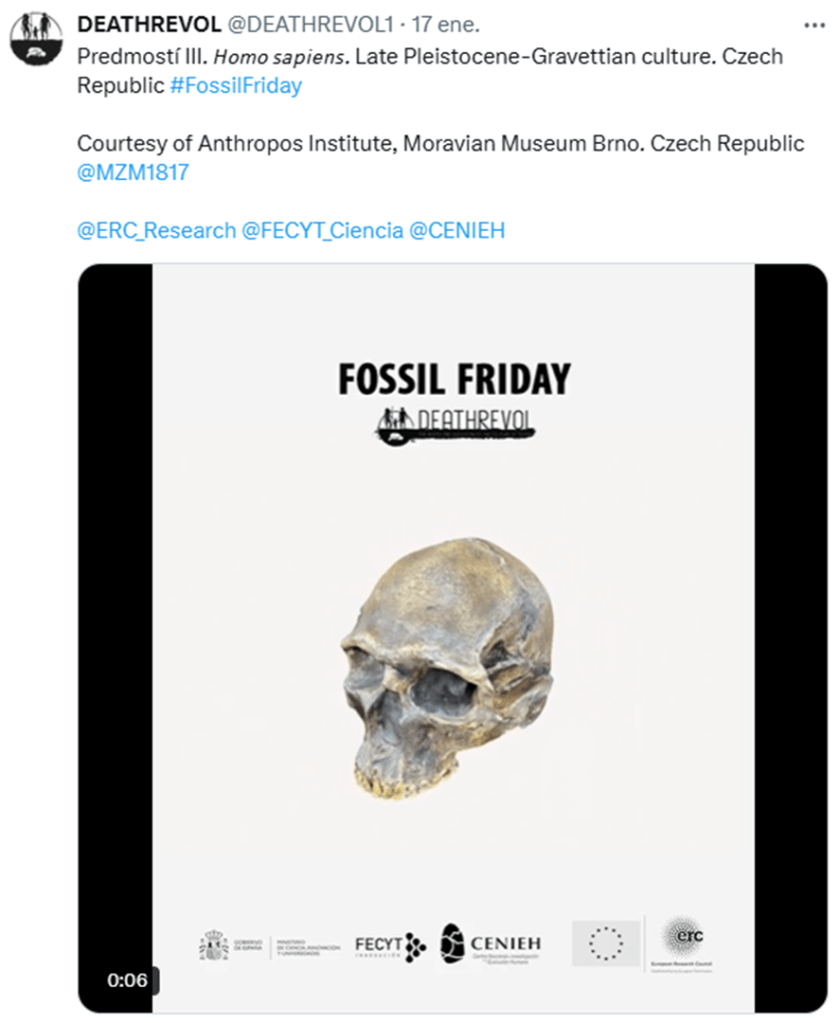
9. Who said fossil skulls can’t be beautiful? By DEATHREVOL @DEATHREVOL1