Poco a poco vamos completando la historia de lo que ocurrió en la cueva Denisova desde hace 300 ka (miles de años). Jacobs, Zavala, Li y otros compañeros describen de forma exhaustiva en su publicación la cronología de los homininos y la fauna que ocuparon la cueva en distintos momentos hasta la llegada de los humanos modernos. Este yacimiento es clave para comprender la evolución de los enigmáticos denisovanos, su interacción con los dos homininos con los que convivió y están muy próximos genéticamente (Homo neanderthalensis y Homo sapiens), así como los cambios faunísticos asociados a las fluctuaciones climáticas de este tramo del Pleistoceno.
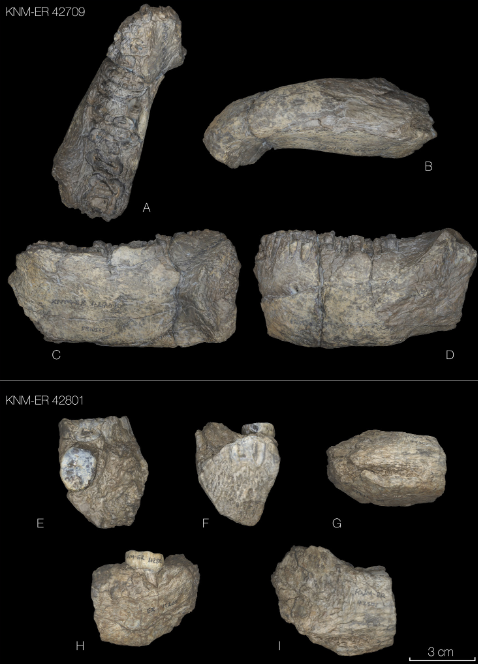
Los hallazgos se basan en dataciones ópticas y el análisis de ADN mitocondrial y de ADN nuclear extraído de sedimentos de las tres cámaras principales de la cueva (Principal, Este y Sur). Por primera vez, las tres cámaras se estudian de manera integral en este trabajo, ya que hasta ahora el foco había estado en la Principal y la Este. En estas dos cámaras se habían hallado hasta ahora seis fósiles de denisovanos, cuatro de neandertales y un fragmento óseo de un descendiente de neandertal-denisovano (Denisova 11), así como fragmentos de ADNmt de denisovanos, neandertales y humanos modernos en 160 muestras de sedimentos. Mientras tanto, en la Cámara Sur se han recuperado dos dientes denisovanos (Denisova 4 y 25), dos fragmentos de cráneo denisovanos (Denisova 13 y 22), y ADNmt hominino en 15 muestras de sedimento.
Sigue leyendo