

¡Hemos cerrado un trimestre cargadito de novedades importantes! Aquí va mi resumen de las 39 seleccionadas:
- Una reevaluación de la morfología de los huesos coxales en el Australopithecus prometheus StW 573 de Sterkfontein Member 2 (3,67 millones de años, Ma), comparada con la variación de los huesos coxales en humanos y simios actuales, así como en otros homininos del Plioceno y del Pleistoceno, sugiere distintas morfologías y múltiples formas de bipedalismo entre los primeros homininos, y un «refuerzo firme» de la hipótesis de una segunda especie de Australopithecus en Sterkfontein [+].
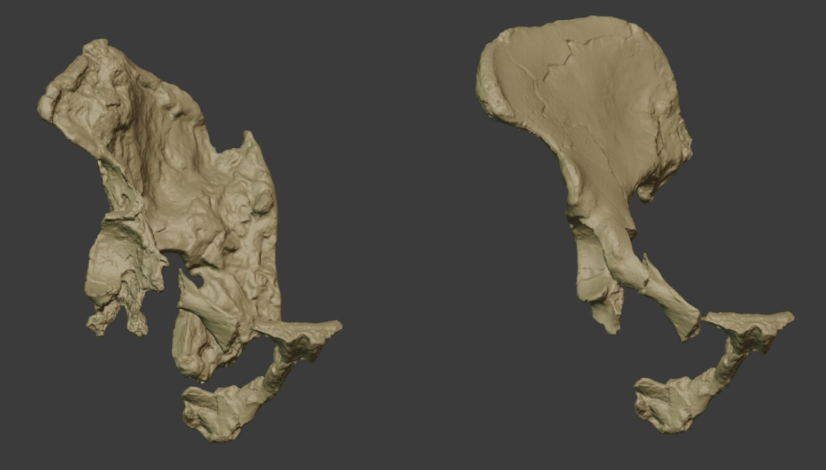
Modelos digitales de StW573 y StW 431. Crédito: R. Crompton et al. (2024).
- Bienvenidos dos nuevos dientes de Paranthropus en Olduvai: los molares OH 92 y OH 30B. Este último procede del sitio Maiko Gully en Frida Leakey Korongo (FLK) y, por su morfología, se ha podido determinar que pertenece al individuo OH 30 hallado en 1969, datado en 1,8-1,7 Ma. En cuanto al OH 92, se halló en el área Bell’s Korongo (BK), pero no se ha podido identificar la capa sedimentaria de donde procede [+].
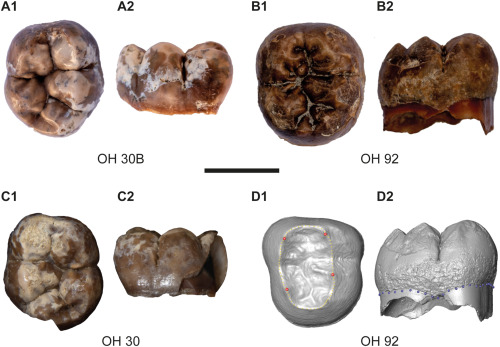
Dientes analizados en A. Riga et al. (2024)
- La comparación entre el conjunto achelense de MW5 (Melka Wakena, Etiopía), de 1,37-1,34 Ma, con otros conjuntos anteriores de 1,6 y 1,7 Ma, muestra una mayor planificación previa para la adquisición de materia prima y una mayor inversión en el control de las propiedades morfométricas de los artefactos, que marcan el cambio hacia una tecnología basada en lascas grandes (Large Cutting Tools, LCT) [+].
- Se ha revisado la datación de los tres yacimientos de Orce (sur de España) que cuentan con algunas de las evidencias más antiguas de homininos en Europa occidental: Venta Micena (1,32 Ma), Barranco León (1,28 Ma) y Fuente Nueva 3 (1,23 Ma) [+].
- Y seguimos en Orce. Aunque se publicó al final del anterior trimestre, no quiero dejar de mencionar el estudio sobre la interpretación de los restos de megafauna hallados en Fuente Nueva 3. La capa 5 del nivel superior tiene suelos de arenas finas y blandas con presencia de muchos restos de megahervíboros, como Mammuthus meridionalis, que serían consumidos por carroñeros como hienas gigantes (Pachycrocuta brevirostris). En las capas 2-3 del nivel inferior, formadas por limos y arcillas, se han hallado numerosos manuports, piedras no modificadas utilizadas por los homininos como herramientas de percusión para fracturar huesos y acceder al tuétano [+].
- Barranc de la Boella puede representar una dispersión temprana del Achelense desde África hace unos 1,4 Ma, potencialmente conectada a conjuntos como ‘Ubeidiya. Este trabajo subraya la complejidad de la tecnología lítica durante la aparición de nuevos comportamientos en los conjuntos europeos de Modo 2 inicial, que se extienden más allá de la mera aparición de LCTs [+].
- La caza humana fue un factor clave en la pérdida de docenas de especies de proboscídeos: la tasa de extinción de estos animales se quintuplicó cuando aparecieron los primeros humanos hace unos 1,8 Ma, y aumentó aún más cuando aparecieron los humanos modernos, según el análisis de miles de fósiles asistido por IA [+]. Hoy en día, solo quedan tres especies de este grupo.
- La concentración de yacimientos en puntos calientes como el Sistema del Rift de África oriental es un factor a tener en cuenta para evitar el sesgo en nuestra comprensión de la evolución humana. Es una discusión que surge de haber estudiado los mamíferos modernos de cuerpo mediano y grande especialistas del Rift, entorno que representa sólo el 1,6% del rango total de los mamíferos africanos. En particular, los cráneos de primates modernos del Rift suponen menos del 50% de la variación total entre todos los cráneos de primates africanos [+].
- El genoma humano de alta cobertura más antiguo hasta la fecha es el de Denisova 25, un denisovano de hace 200 miles de años (ka), 80 ka más antiguo que el genoma del neandertal de la cueva siberiana de Chagyrskaya. Se trata de un individuo masculino que proviene de una población distinta de denisovanos tempranos que se cruzaron varias veces con un grupo de neandertales cuya población no había sido detectada antes en el ADN. De ésta heredó el 5% de su ADN. Es el segundo genoma denisovano secuenciado, tras el de la falange distal de Denisova 3 (60-80 ka). Todavía no publicado, este trabajo se anunció en la reunión anual de la Society for Molecular Biology and Evolution [+].
- Utilizando genomas de 2000 humanos modernos, así como de tres neandertales y un denisovano, se ha conseguido mapear el flujo genético recurrente entre los tres grupos humanos durante los últimos 200 ka: una primera oleada de contactos hace unos 200-250 ka, otra hace unos 100-120 ka y la mayor hace unos 50-60 ka [+].
- Un fragmento de húmero de 700 ka hallado en Mata Menge (Indonesia) procede de un individuo adulto que mediría unos 100 cm, es decir, unos 6 cm menos que la estatura estimada según la longitud del fémur para el esqueleto de Homo floresiensis de Liang Bua, de 60 ka. De hecho, es el húmero más corto del registro fósil hominino [+]. Recordemos que los molares de Mata Menge también son más pequeños que los conocidos de cualquier otro hominino.
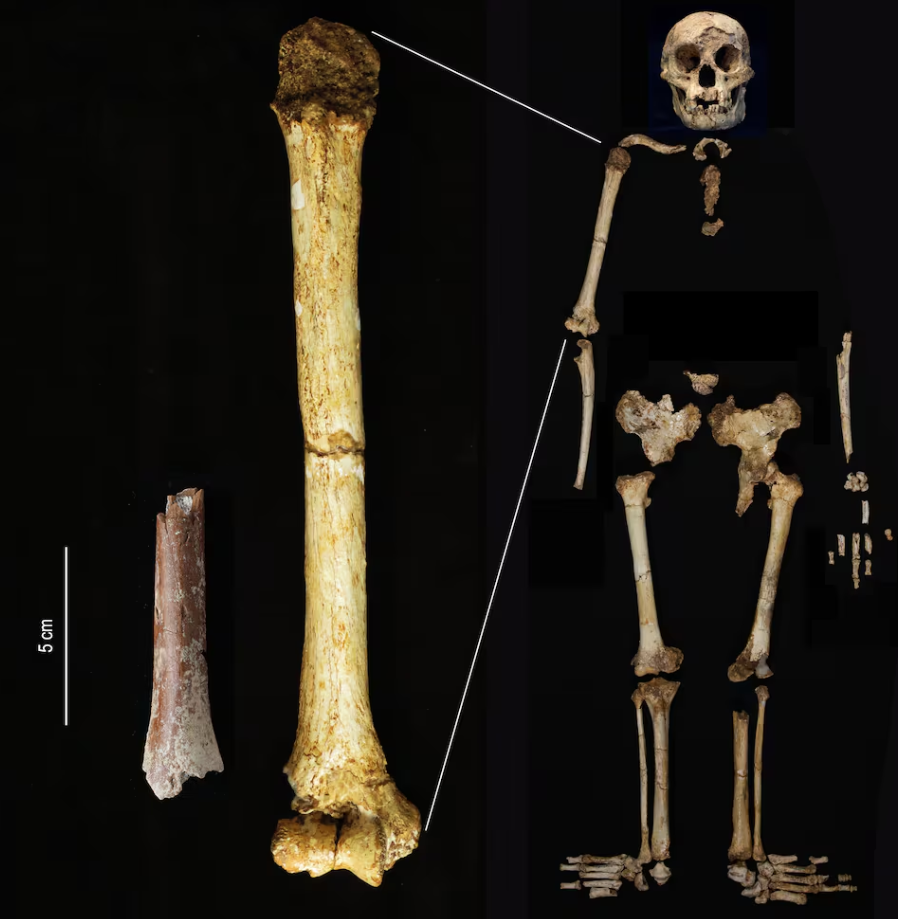
Fragmento de húmero de Mata Menge (izda) a la misma escala que el húmero de Homo floresiensis de Liang Bua (centro y derecha). Crédito: Yousuke Kaifu.

