Me había propuesto en este trimestre ser realmente estricto en el número de novedades seleccionadas. Os aseguro que lo he sido, a pesar de lo cual han sido 49 las que recopilo. Estamos en un tiempo fantástico para la paleoantropología.
- Las tasas de cicatrización de heridas humanas son tres veces más lentas que en primates no humanos, las cuales, a su vez, son similares a las de otras especies animales como roedores. Matsumoto-Oda y colaboradores se plantean la desventaja evolutiva que pudo haber supuesto la pérdida del vello corporal. A diferencia de los grandes simios, los humanos experimentaron en su evolución un aumento en la densidad de las glándulas sudoríparas y una disminución en la densidad del vello corporal, acompañado del engrosamiento del tejido subcutáneo como forma alternativa de protección interna. Esto habría contribuido a la evolución de tasas de cicatrización más lentas, que pueden dificultar el acceso a los alimentos, reducir la capacidad de evadir depredadores e influir sobre las necesidades de energía para la reparación de tejidos, el crecimiento y la reproducción. Es posible que el desarrollo del cuidado a individuos mayores y discapacitados entre humanos, así como el uso de plantas medicinales, mitigaran la desventaja evolutiva de la cicatrización tardía de las heridas.
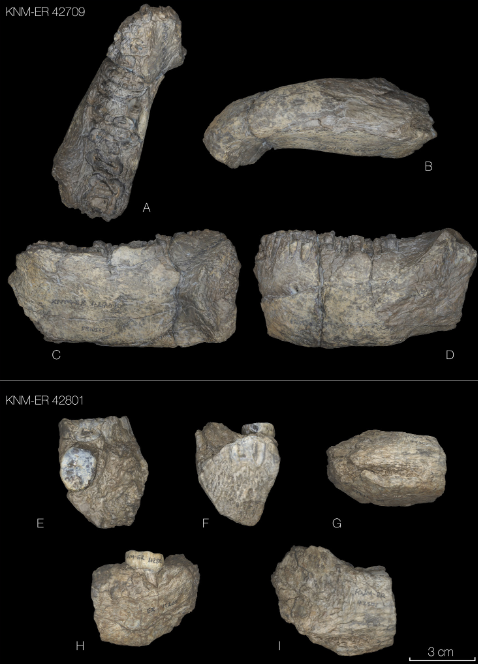
- Los primeros Homo (incluyendo H. habilis y H. rudolfensis) y Paranthropus boisei son propuestos como los fabricantes de útiles olduvayenses (Williams et al., 2025). Los primeros miembros del género Homo siguen siendo los principales candidatos para la fabricación y el uso del Olduvayense a lo largo de su rango temporal más amplio (c. 3,25-1,6 Ma, millones de años) pero, considerando que el Olduvayense pudo darse hasta hace 1,2 Ma, P. boisei sería el principal candidato para la producción de los artefactos posteriores.

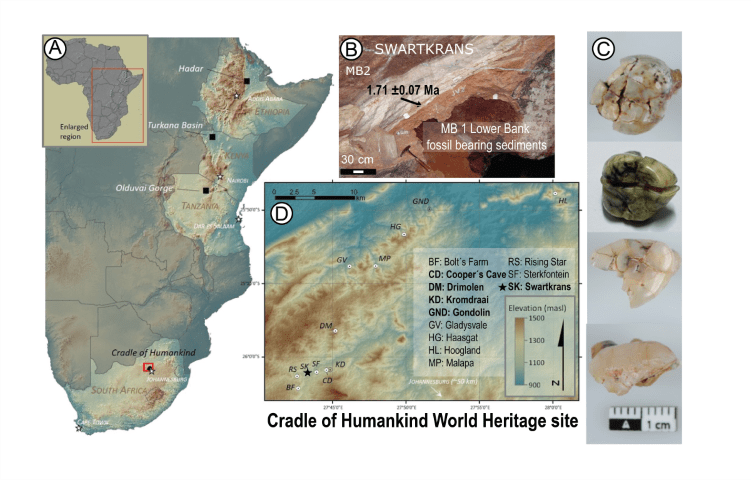
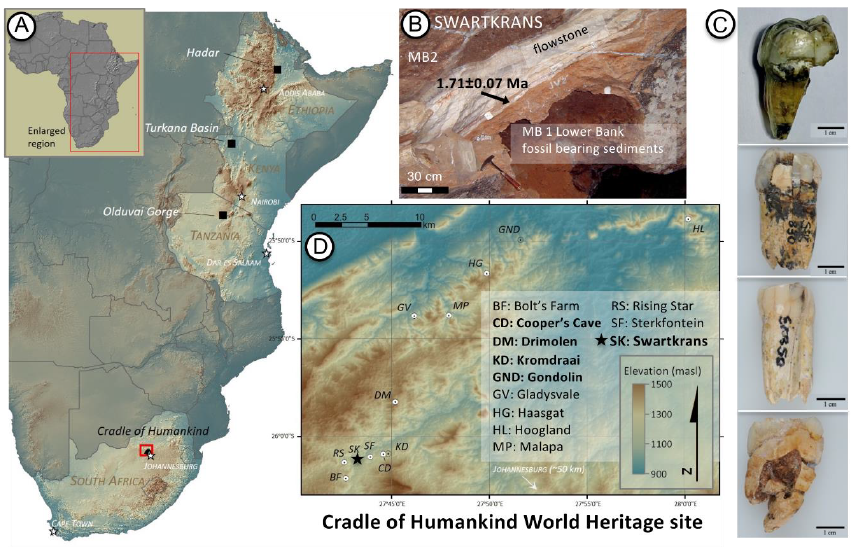
- El espécimen SWT1/HR-2 de Paranthropus robustus, compuesto por un os coxae parcial articulado, el fémur casi completo y la tibia completa, procedente de Swartkrans Miembro 1 (1,7-2,3 Ma), indica bipedación completa y una estatura de solo 1,03 m, el hominino más pequeño conocido (Pickering et al., 2025).
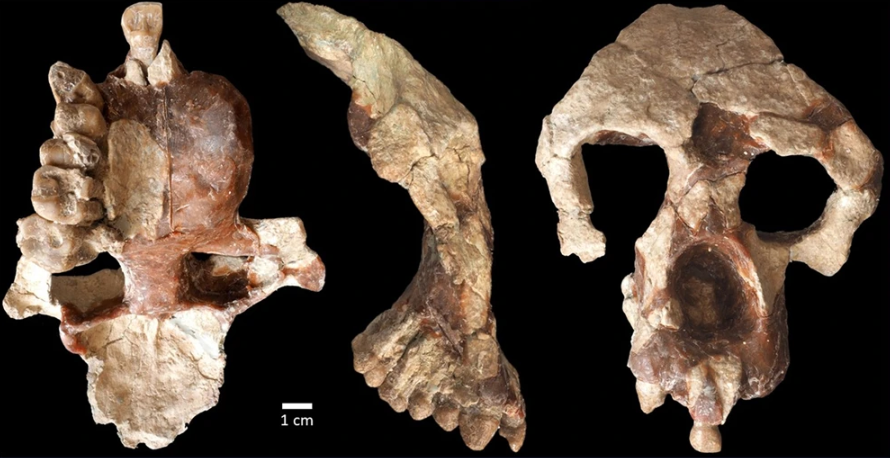
- Tras conocer el sexo biológico del espécimen Sts 63 (Australopithecus africanus) del Miembro 4 de Sterkfontein gracias a su proteoma conservado en un diente (Madupe et al., 2025), ahora han seguido el mismo camino cuatro especímenes de Paranthropus robustus de Swartkrans, de unos 2 Ma: son los dientes SK 830, SK 835, SK 850 y SK 14132. El estudio de la proteína amelogenina ha podido determinar que dos de los dientes tenían la variante específica de los machos (Madupe et al., 2025). Además, el polimorfismo de un solo aminoácido y la variación en la forma de la unión esmalte-dentina indicaron subgrupos potenciales presentes dentro de P. robustus. Anteriormente, se tenía identificada una variante genética única para la proteína polishina en dicha especie, pero este estudio mostró variaciones entre los cuatro individuos analizados. Esto podría reflejar dinámicas poblacionales complejas, microevolución o la presencia de grupos o taxones distintos.