Este trimestre que cerramos ha venido cargado de novedades: nada menos que 49 trabajos recopilados, que muestran cómo este campo sigue despertando interés, tanto en la comunidad científica como en el público general. Ahora bien, siguiendo mi campaña contra los titulares exagerados, conviene recordar que cada hallazgo no supone “reescribir la historia de la evolución humana”. ¿Lo habríamos tenido que hacer 49 veces en un solo trimestre? Comprendamos el significado de cada avance en su contexto.
- El ilion experimentó dos innovaciones clave en la evolución humana que hicieron posible la bipedación: la reorientación de la placa de crecimiento ilíaca perpendicular a la columna vertebral, y una osificación ilíaca humana tardía (Senevirathne, Fernandopulle, Richard et al., 2025).
- Los chimpancés del valle de Issa (Tanzania) pasan tanto tiempo en los árboles como los chimpancés de bosques más densos. Además, no solo trepan, sino que también suelen ponerse de pie y desplazarse erguidos sobre las ramas, usando los brazos para equilibrarse mientras buscan alimento. Se trata de un entorno parecido al de los primeros homininos de hace entre 4 y 7 Ma (millones de años), que aún mostraban adaptaciones claras para trepar, como brazos largos y dedos curvados. Aquellos homininos pudieron haber combinado vida arbórea y terrestre y desarrollado el caminar erguido en las ramas, aprendiendo el equilibrio antes de dominar la marcha en espacios abiertos (Drummond-Clarke et al., 2025).
- Baker, Barton y Venditti han hallado en 95 primates fósiles y actuales una correlación entre la longitud del pulgar, una característica clave del agarre de precisión, y el tamaño del cerebro, utilizando métodos filogenéticos bayesianos. En concreto, no se ha encontrado un vínculo con el tamaño del cerebelo, pero sí una fuerte relación con el tamaño del neocórtex, lo que puede reflejar el papel de las cortezas motora y parietal en las habilidades sensomotoras asociadas con la manipulación fina.
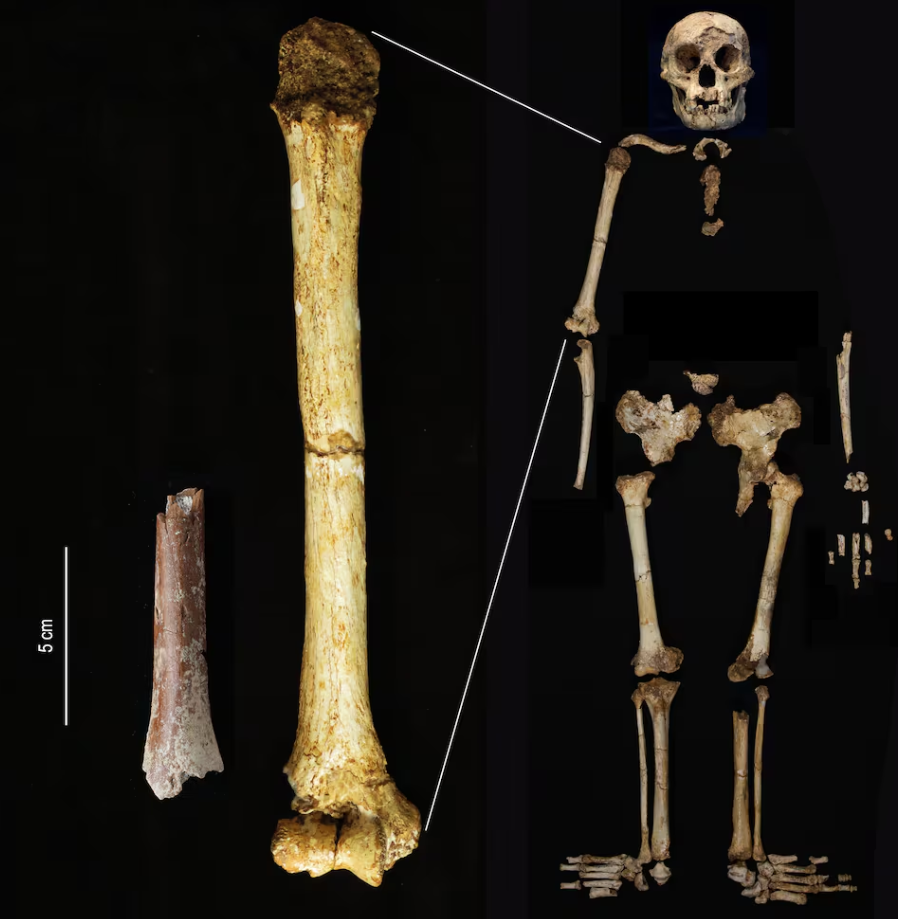
- El análisis de ocho dimensiones poscraneales sugiere que tanto Australopithecus afarensis como Australopithecus africanus presentan un alto grado de dimorfismo sexual de tamaño, significativamente mayor que el de los chimpancés y los humanos modernos. A su vez, Au. afarensis tiene mayor dimorfismo que Au. africanus (Gordon, 2025).
- El estudio de las clavículas de Au. afarensis, en particular las del individuo juvenil DIK-1-1 y del adulto KSD-VP-1/1, en comparación con las de simios actuales, revela una geometría cortical que sugiere un comportamiento arbóreo sostenido en estos homininos, incluyendo suspensión y escalada (Farrell y Alemseged, 2025).
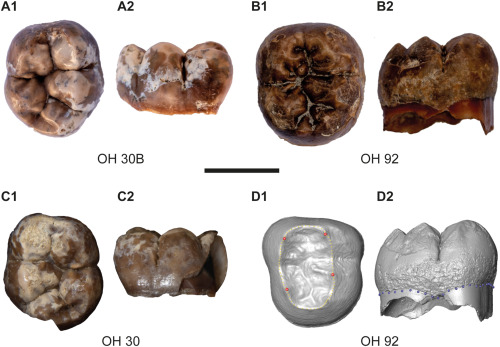
- La introducción en la dieta de los australopitecos de vegetales duros como las gramíneas precedió en unos 700 ka a la adaptación de su dentición, con el desarrollo de molares más largos y la reducción del tamaño de los dientes en un 5% cada 1000 años. Fannin y colaboradores han observado que los Au. afarensis dejaron de consumir frutas, flores e insectos para alimentarse de pastos y juncos hace entre 3,4 y 4,8 Ma, a pesar de carecer de los dientes y el sistema digestivo óptimos para comer estas plantas más duras y resistentes. Los homininos habrían podido maximizar los nutrientes derivados de estas plantas a medida que sus dientes se volvían más eficientes para descomponerlas. Posteriormente, hace unos 2 Ma los isótopos de carbono y oxígeno en los dientes de los primeros Homo cambiaron abruptamente, por la reducción del consumo de pastos y el aumento relativo del agua probablemente contenida en tubérculos, bulbos y cormos, tejidos vegetales de alta energía. En este momento los dientes muestran un cambio repentino en forma y tamaño más adecuado para comer tejidos cocinados.
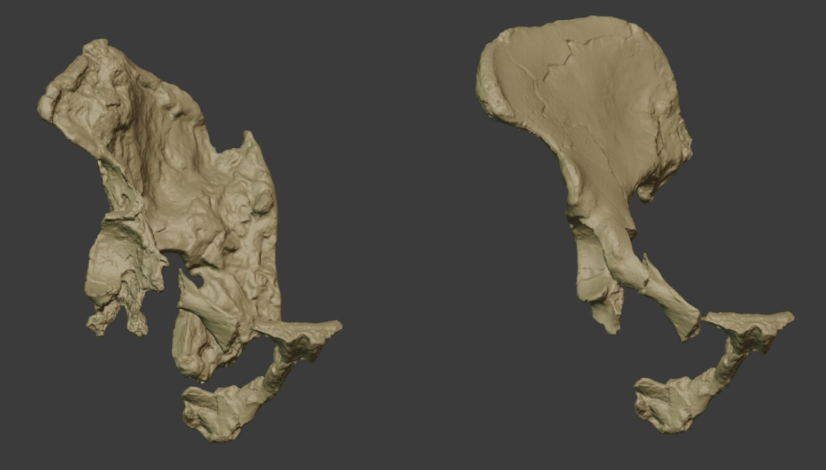
- Los homininos de Nyayanga (Kenia) fabricaron herramientas olduvayenses hace 2,6 Ma principalmente a partir de materia prima no local, transportada hasta 13 km para aprovechar su calidad (Finestone et al., 2025). Con esta antigüedad, los autores de aquellas herramientas pudieron ser representantes de Homo tempranos o Paranthropus, cuya presencia se ha documentado en el mismo nivel NY-1. Las anteriores evidencias de transporte de materias primas estaban datadas en unos 2 Ma, procedentes de Kanjera South (a 15 km de Nyayanga) y de Olduvai y Ewass Oldupa en Tanzania. El mismo trabajo recopila una comparativa de lugares y distancias de transporte. Se han observado transportes similares realizados por chimpancés durante unos 2 km.