Recientemente tuve la oportunidad de entrevistar a Eduardo Méndez Quintas, investigador principal del conjunto de LCTs (large cutting tools) del yacimiento de Porto Maior (sobre este sitio escribí recientemente aquí). Eduardo es experto en el estudio del origen de la industria achelense en África y de su extensión por Europa, periodos apasionantes en la evolución humana. En relación con este tema, nos comparte para este blog las últimas novedades en su investigación actual.
En 2018 tuvo gran repercusión vuestra publicación en Scientific Reports sobre las LCTs de Porto Maior. ¿Cuál es la importancia de este trabajo?
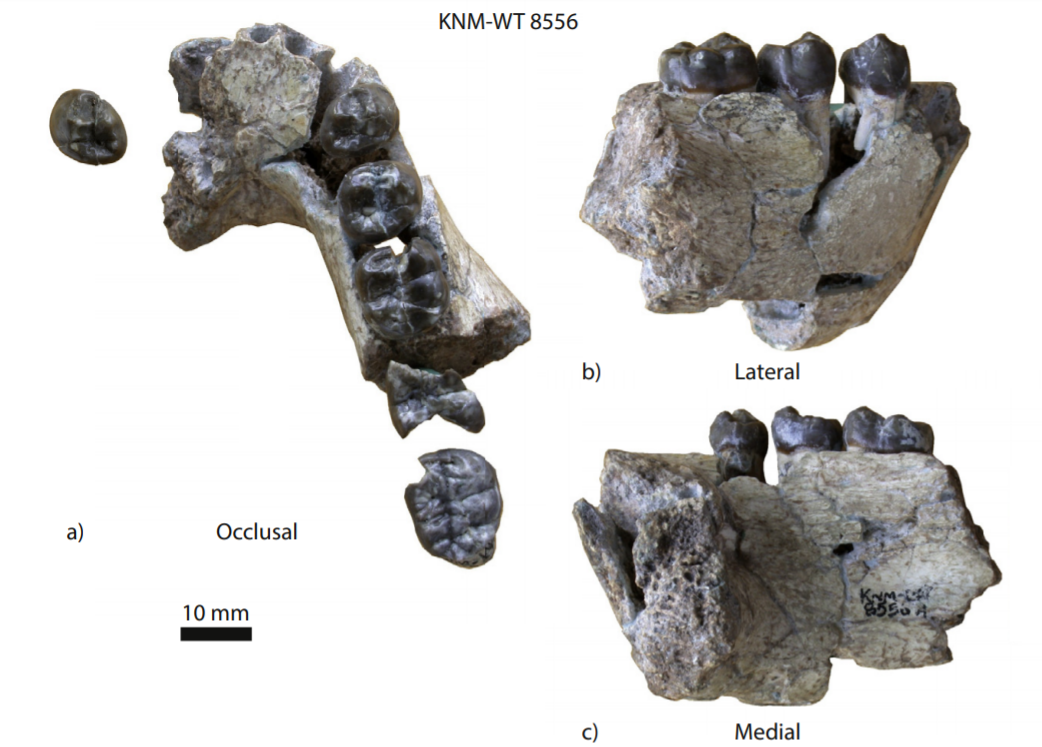
Porto Maior tiene fundamentalmente dos niveles con evidencias de ocupación humana: el PM3 y el PM4. Por encima de estos, hay un depósito muy reciente, del Último Máximo Glacial, que tiene material retrabajado, que viene arrastrado y erosionado. El más conocido es PM4 porque tiene el conjunto de LCTs (Large Cutting Tools). Se trata de un tapiz increíble de piezas, con 10-12 bifaces por m2, en los 40 m2 que hemos excavado. Son materiales con talla muy parecida: muestran gestos que son iguales, pero también los bifaces resultantes son idénticos en varios casos. Pensamos que pueden estar hechos en un periodo relativamente corto y por un número limitado de personas.
Lo más interesante es que casi todas las piezas están acabadas. Las lascas y núcleos que se encuentran son material que está rodado, viene arrastrado con el sedimento. Lo que vemos es una acumulación de piezas acabadas, una especialización. Las piezas en sí mismas son aparentemente toscas y elementales, con un nivel de configuración bajo: pocos levantamientos, grandes y hechos con percutor duro. Fuera de África, salvo en Gesher (Israel), no hay ningún otro conjunto con una acumulación tal de LCTs achelense.
En África los yacimientos con grandes acumulaciones de bifaces son normalmente excluyentes con la presencia de fauna. Por ejemplo, colaboramos en la investigación del yacimiento Garba I en Melka Kunture (Etiopía), que fue excavado por Jean Chavaillon en los años 1960. Tiene una antigüedad de ~600 miles de años (ka) y en 240 m2 aparecieron más de 7000 herramientas, sobre todo bifaces. En este yacimiento se conservan restos de hipopótamo y otra fauna, pero son muy minoritarios en comparación con la industria lítica. Otro ejemplo lo tenemos en el sitio TK, que está en el Lecho II de Olduvai y tiene 1,3 millones de años (Ma): en el nivel llamado Lower Ocupation hay una acumulación muy densa de grandes configurados, más grandes que los de Porto Maior, mientras que en el nivel Upper Occupation hay restos de fauna (elefante, Sivaterio, etc.) junto con bifaces, pero éstos ya son de un tamaño más «normal».
Si los bifaces no se empleaban para el procesado de animales, ¿cuál pensáis que era el objetivo de esta acumulación?
Vamos a realizar estudios de traceología para intentar buscar respuestas. En los primeros que hemos iniciado, ya hemos visto que muchas piezas muestran un uso que sería compatible con el procesado de materiales duros, como la madera y el hueso, lo que nos hace pensar en el aprovechamiento de estos materiales.
Otra cuestión interesante a estudiar es que los bifaces no aparecen de forma homogénea en el sedimento, sino agrupados por clusters. En África, muchas veces los yacimientos tienen procesos sedimentarios que tienden a acumular las piezas. Aquí no tenemos un medio tractivo, así que las piezas están más o menos como se dejaron, lo cual ayuda a estudiar el trasfondo de ese comportamiento y utilización del espacio.
Hay quien propone una explicación para la acumulación de piezas en sitios africanos, planteando que las herramientas de formatos grandes y apuntados ayudarían para acceder a los conductos de termiteros en el suelo. Pero esta hipótesis es difícil de plantear en Europa, ya que no consta la existencia de termiteros en el Pleistoceno medio europeo. Otra idea de Richard Potts propone que los homininos usarían escondrijos en determinados puntos del paisaje africano, como remanente de piezas finalizadas para ser utilizadas en sus desplazamientos.
En cualquier caso, primero debemos validar la presencia de huellas de uso en todas las herramientas y, si el resultado es positivo, plantearnos el motivo. La hipótesis del taller para el procesado de madera podría funcionar, ya que conocemos herramientas en este tipo de material casos de esa antigüedad (Schöningen, Clacton…), que son puntuales porque la madera apenas se conserva, pero estamos seguros de que sería un material usado muy habitualmente.

Foto: LCT de Porto Maior PM4. Crédito: Roberto Sáez
Sigue leyendo →