La bipedación es una forma única de desplazamiento empleada por una sola de todas las especies de primates actuales, y se considera uno de los rasgos esenciales que define al linaje humano. Por ello, intentar conocer cuándo, dónde y por qué surgió esta modalidad de locomoción suele generar un rico debate.
El Mioceno, periodo entre hace 23 y 5 millones de años (Ma) en ocasiones suele llamarse «planeta de los simios», por las numerosas especies de primates que poblaban entonces Europa, África y Asia: las conocidas se cuentan por decenas, y cuántas otras existirían que nunca conoceremos… Esas especies nos van aportando datos del camino evolutivo que siguieron los primates, pero el registro fósil no es abundante y hay muchos huecos, por ejemplo, a partir de hace 7 Ma hasta la aparición de los australopitecos (4 Ma). Por ello, todavía es muy difícil comprender la transición hacia el bipedismo desde otros hábitos (saltación, suspensión, cuadrupedismo arbóreo, cuadrupedismo terrestre), o averiguar cuál sería el comportamiento locomotor del último ancestro común de humanos y chimpancés.
En los primeros homininos vemos ciertos indicios de haber desarrollado una postura erguida, pero no un desarrollo especializado de la bipedación como sí se observa posteriormente:
- La posición del foramen magnum de Sahelanthropus tchadensis manifiesta una cierta posición erguida (en espera de conocer un misterioso fémur de esta especie, que parece existir pero no se publica).
- Orrorin tugenensis presenta menos dudas de poder mantener una posición erguida, aunque contamos con muy pocos huesos.
- Ya hay rasgos más claros en la morfología de distintas partes del esqueleto de los ardipitecos, de que eran seres bípedos en ambientes boscosos hace más de 4 Ma, pero también hábiles trepadores.
- Finalmente, los australopitecos serán los primeros seres que muestran claramente una locomoción bípeda habitual hace entre 4-2,5 Ma, manteniendo aún ciertas capacidades de trepa. De entre estos seres posiblemente surgió el linaje humano.
Pero todo esto no significa que la bipedación sea exclusiva de los humanos sino que, dentro de otras especies de primates anteriores a los homininos, pudieron ir apareciendo «pruebas» de bipedación y de otros hábitos locomotores distintos a los que empleaban, como adaptaciones de su morfología más adecuadas a su modo de vida.
Así, este artículo resume dos recientes estudios que aportan nuevos datos muy interesantes sobre este tema.
Oligoceno (23-34 Ma): Aegyptopithecus
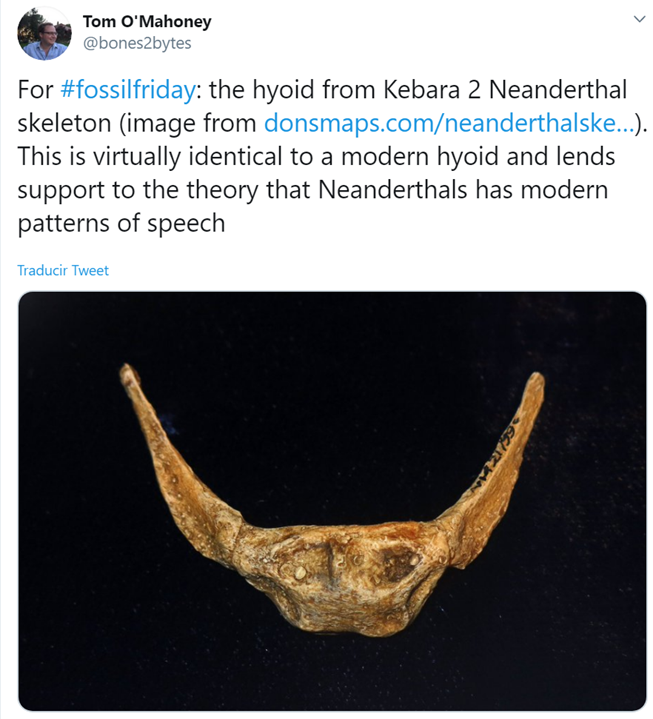
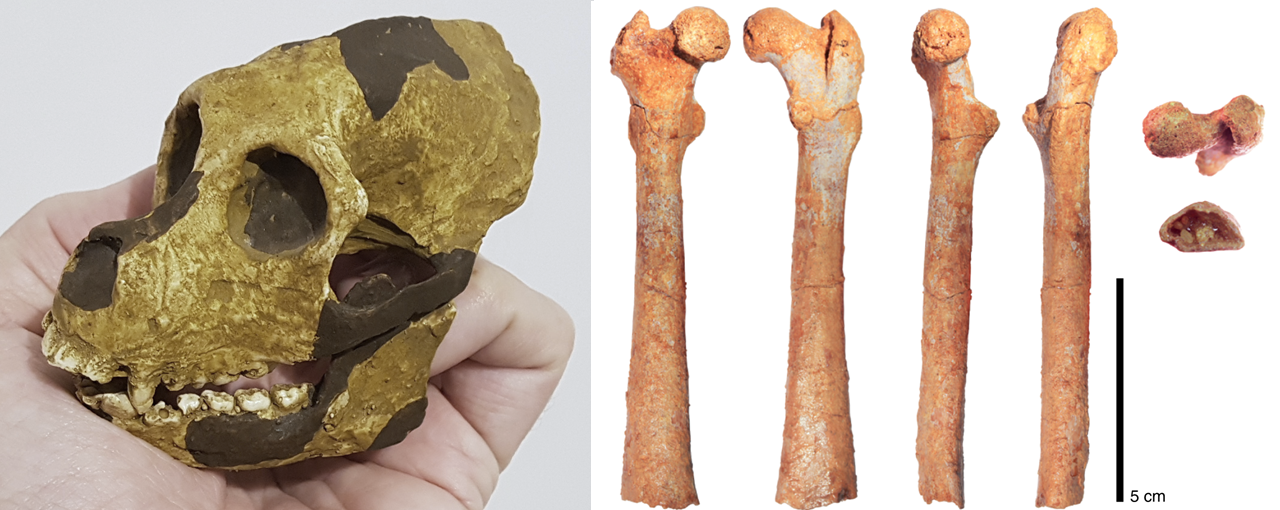
Este es el cráneo de Aegyptopithecus zeuxis, que vivió en un momento previo a la separación de las dos superfamilias de catarrinos: los hominoideos (gibones, orangutanes, gorilas, chimpancés, humanos) y los cercopitécidos (monos del Viejo Mundo: babuinos, cercopitecos, macacos, colobos).

1) Reproducción de cráneo de A. zeuxis basada en dos fósiles (AMNH 13389 y YPM 21032) encontrados en El Fayum (Egipto) – crédito: Roberto Sáez. 2) Anterior, posterior, medial, lateral, proximal and distal views, respectively, of DPC 24466 – credit: Almécija S. et al. (2019)
Sigue leyendo →