[English version below]
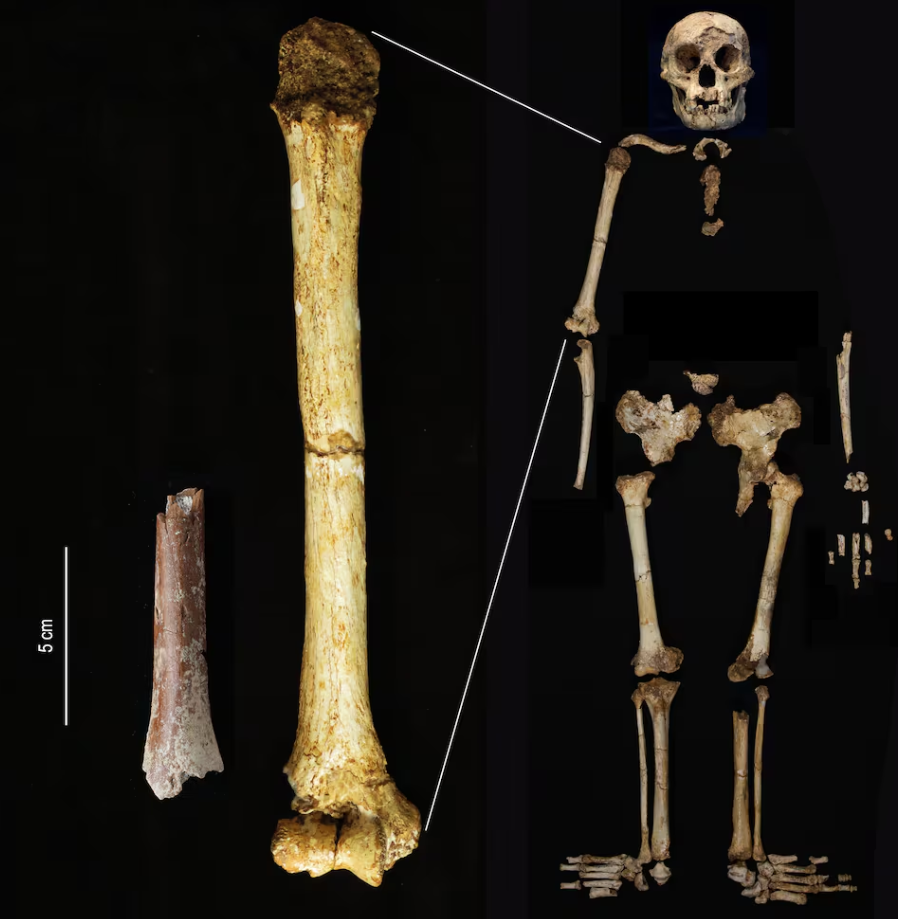
A orillas del lago Turkana, en Kenia, una muchacha vivió meses postrada a causa de una grave enfermedad que le provocaba dolores en abdomen y cabeza, mareos, visión borrosa, pérdida de coordinación muscular… Se le fue cayendo el pelo, la piel se le agrietaba y sufría hemorragias internas. Por dicha, tuvo a su lado a alguien (cuya identidad desconocemos) para cuidar de ella día y noche, acercarle agua y comida, protegerle y acompañarle hasta su fallecimiento.
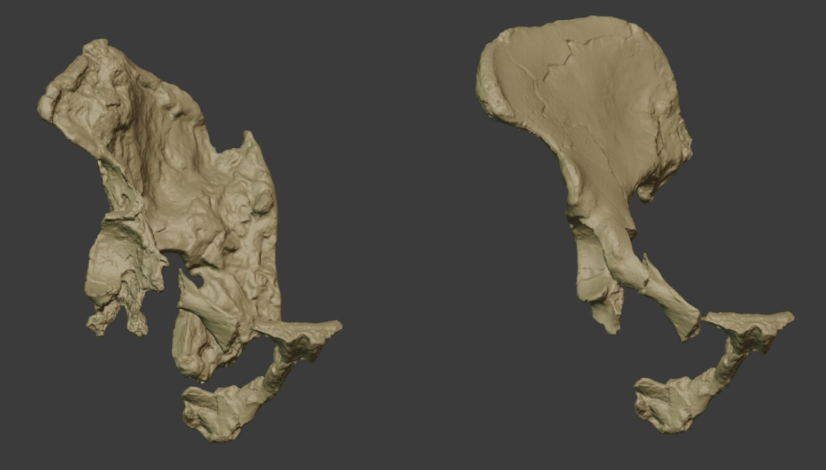
Esta situación parece una de tantas en que personas atienden a personas, forma parte de nuestra vida cotidiana. Lo singular del caso es que sucedió hace casi dos millones de años. El estudio de los fósiles encontrados en el yacimiento de Koobi Fora ha permitido a los arqueólogos reconstruir la patología de esa mujer de la especie Homo ergaster que solo pudo sobrevivir algún tiempo con ayuda. Sus huesos son un testimonio conmovedor de los inicios de la sociabilidad entre nuestros ancestros homínidos.
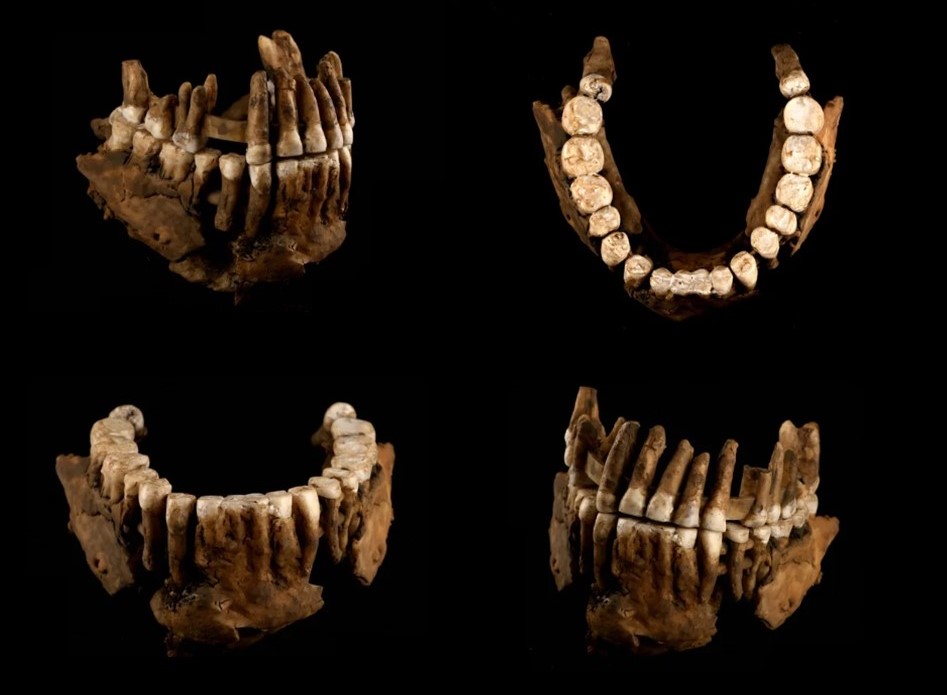
Decía Ortega que estar abierto al Otro es un estado permanente y constitutivo del ser humano. Pero ¿es una cualidad exclusivamente humana? Porque la Red rebosa de vídeos y de testimonios de animales que colaboran entre sí, que socorren a congéneres en apuros o que parecen sumirse en duelo por sus amos. ¿Cuándo empezó a manifestarse en la historia la compasión, el altruismo y el mutuo cuidado como comportamiento sistémico de nuestra especie? Hasta hace poco ello se asociaba a los modernos, a nuestro camino evolutivo más reciente, pero casos como el citado de la muchacha de Koobi Fora o el de ‘Benjamina’, la niña discapacitada de Atapuerca (cuyo cráneo ha sido definido como «la primera muestra de amor fosilizado»), revelan que esa intencionalidad viene de épocas muy lejanas.
Muchos científicos piensan que la identificación emocional tuvo un papel determinante en el desarrollo de comportamientos humanos y en nuestros modos de vida. Lejos de hacemos animales más vulnerables, esa capacidad permitió al grupo superar las dificultades y salir adelante. Por añadidura, la reciprocidad social proporciona satisfacción, lo que favorece la selección de genes que nos inclinan a cooperar y a la apertura al Otro, De manera que, desarrollando la empatía, experimentamos el placer de descubrir el mundo mental de los demás.
Todo lo anterior lo explica con claridad y en síntesis el ingeniero y divulgador científico Roberto Sáez en un libro de reciente aparición titulado ‘Evolución humana: Prehistoria y origen de la compasión’ (editorial Almuzara). Encandilante y conmovedor.
¿Cuándo empezó a manifestarse en la historia la compasión, el altruismo y el mutuo cuidado como comportamiento sistémico de nuestra especie?
Por Juan Aguirre. Artículo publicado en EL DIARIO VASCO  Sigue leyendo →
Sigue leyendo →