Cerramos 2021 con 24 novedades en paleoantropología publicadas en este último trimestre, que recopilo por orden cronológico. En total han sido ¡112 noticias! las que he ido incluyendo en mis resúmenes trimestrales durante este año. Y creedme que he sufrido bastante para poder seleccionar.
-
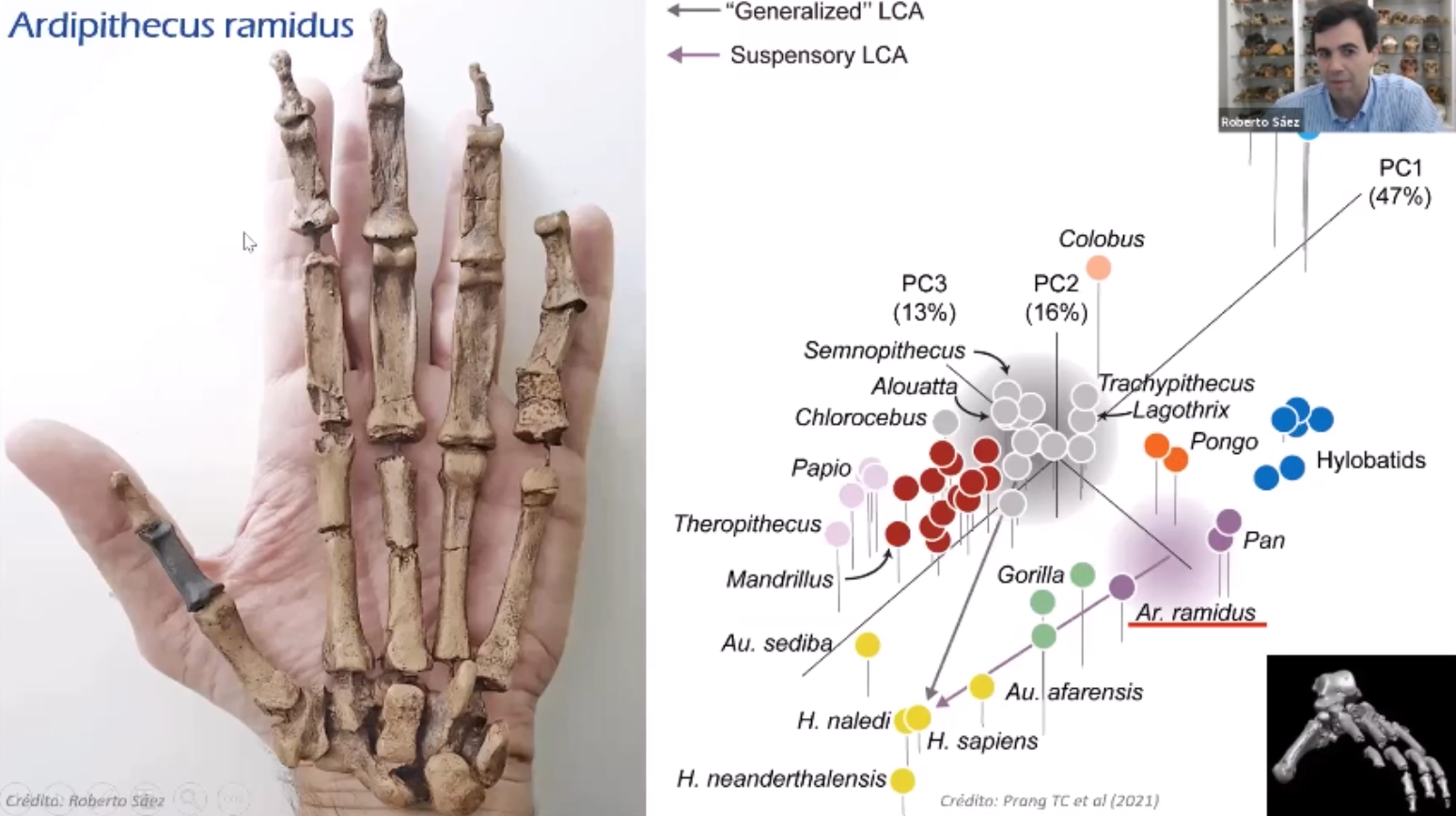
Se ha definido un nuevo método para determinar el nivel de dimorfismo sexual de los dientes caninos de forma fiable en simios antropoides extinguidos. Este estudio concluye que el dimorfismo sexual de los caninos era probablemente muy bajo en los homininos ya desde Ardipithecus ramidus, hace 4,5 Ma (millones de años), así como en las especies posteriores de Australopithecus y Homo, prácticamente indistinguible al de los humanos modernos, y muy inferior al del bonobo, que es el menos dimórfico y conductualmente menos agresivo de los grandes simios actuales [+info]. Puede que todos tuviéramos más o menos este concepto en mente, pero nuevos trabajos como este con el método pdPeak (pico de densidad posterior) vienen a confirmarlo.
- Se ha reestudiado un rastro de 5 huellas dejadas en Laetoli (sitio A) hace 3,66 Ma, descubiertas en 1976, dos años antes que el famoso rastro (sitio G) que ha perdurado en todos los textos. Aunque en su momento las del sitio A quedaron olvidadas por las dudas que despertaban sobre el tipo de animal que las hizo (tal vez un oso), el nuevo análisis concluye que los autores fueron homininos, pero distintos a los Australopithecus afarensis a los que se asocian los rastros de los sitios G y S, hallados en 1978 y 2014 respectivamente [+info].

Huellas de Laetoli, sector A. Crédito: Austin C. Hill y Catherine Miller.