Hace tiempo que descartamos una supuesta linealidad evolutiva entre Homo habilis y Homo erectus en los comienzos de «lo humano». Por una parte, tenemos habilinos cada vez más recientes, como el maxilar KNM-ER 42703, de tan solo 1,44 Ma (millones de años) y, por otra parte, tenemos erectinos cada vez más antiguos, como el cráneo parcial DNH 134 de Drimolen, Sudáfrica (1,95-2,04 Ma), y el fragmento de occipital KNM-ER 2598 (1,9 Ma). Donde antes los textos ilustraban una posible evolución anagenética desde unos seres más pequeños y primitivos hasta otros con un cerebro y un cuerpo más grandes y unas piernas más largas, ahora vamos intuyendo un cada vez más probable ancestro común de ambos linajes, todavía por conocer, que dio lugar a la gran diversidad de formas africanas cuya interpretación nos trae de cabeza. De hecho, este antepasado podría haber tenido más rasgos habilinos que erectinos, si atendemos a los pequeños cráneos de Dmanisi (Georgia), de 1,8 Ma. En este otro artículo escribí sobre ello en detalle.
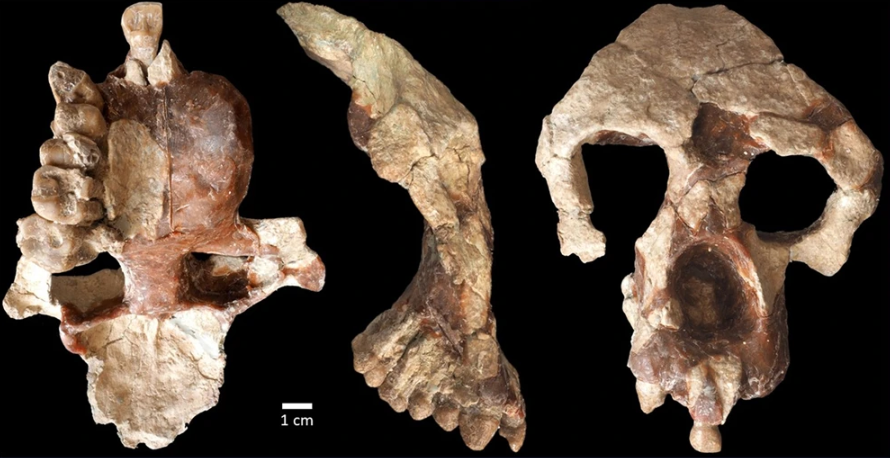
Esta semana hemos conocido a un fósil que se une a la lista de los erectinos más antiguos. La mandíbula infantil Garba IVE (apodada simplemente Garba) se halló en 1982 en el nivel E del yacimiento Garba IV, uno de los más antiguos del complejo de sitios arqueopaleontológicos de Melka Kunture (Etiopía), que abarca tecnologías olduvayense, achelense y MSA. Esta mandíbula de un niño de 2-3 años se ha reevaluado con la ayuda de escáner MicroCT para analizar en el interior del hueso los dientes definitivos no erupcionados, ya que en el exterior sólo se conservan dos dientes deciduos, poco representativos para la identificación a nivel de especie. La anatomía de estos dientes definitivos confirman la adscripción de Garba al taxón de Homo erectus, que ya se intuía por la morfología que la relacionaba con especímenes subadultos como KNM-ER 820 y KNM-ER 1477. De acuerdo a los últimos trabajos de datación del sitio, Garba se convierte en el representante más antiguo de esta especie, con 2,06 Ma de antigüedad.
Adicionalmente, el trabajo ha permitido identificar artefactos de talla achelense en el nivel D, fechado en 1,95 Ma, adelantando en ~0,2 Ma la primera presencia conocida de esta tecnología (por ejemplo, la del FLK West, de 1,7 Ma). Por su parte, la tecnología presente en los niveles inferiores de Garba, de 2 Ma, es olduvayense.

Herramientas achelenses del Garba IV (1,95 Ma). Mussi et al (2023). Cortesía de Eduardo Méndez Quintas.