The #FossilFriday tradition seems to be losing steam, and even hashtags themselves may no longer be as popular as they once were. This trend is also reflected in the declining number of #FossilFriday posts featuring hominin fossils. Still, as long as there continue to be outstanding contributions like those we’ve seen this year, this Top 10 list will live on!
But what exactly is ‘FossilFriday’? Every Friday, social media users share captivating fossil images under the hashtag #FossilFriday. These posts often showcase iconic specimens, museum collections, scientific papers, blog articles, quirky fossils, and much more. I especially enjoy joining in by posting about hominin fossils. So, without further ado, here’s the Top 10 of 2025. Keep posting!
10. The hypnotizing temporomandibular joint of KNM-ER 23000. By Marta Mirazón Lahr @MartaMLahr
9. Who said fossil skulls can’t be beautiful? By DEATHREVOL @DEATHREVOL1
Acompañando al cierre del segundo trimestre, aquí va el resumen de 32 novedades publicadas. Os deseo buena lectura y un feliz verano (o invierno, en el hemisferio sur).
Damos la bienvenida a un nuevo homínido, denominado Buronius manfredschmidi, el más pequeño conocido (10 kg de masa corporal). Datado en 11,62 Ma (millones de años), procede de Hammerschmiede (Bavaria, Alemania). Su holotipo es un molar M2 y paratipos un premolar P4 y un fragmento de paladar. Contrastando con la frecuente simpatría de hominoides (más de una especie conviviendo en el mismo área) en el Mioceno inicial y medio de África, hasta ahora no se conocía ningún caso de simpatría en el Mioceno europeo. En el mismo nivel 5 de Hammerschmiede, se halló el Danuvius guggenmosi presentado en 2019.
La morfología mediofacial varía mucho entre los grandes simios y los humanos, en los que la cara es pequeña y retraída. Aún se desconocen en gran medida los procesos de desarrollo subyacentes a estas diferencias morfológicas. El estudio de la cara media de distintos primates concluye que, en general, los humanos comparten el patrón de modelado óseo mediofacial de los grandes simios, pero la reabsorción ósea es elevada en la zona canina desde el nacimiento, lo que puede ser una clave de la reducción canina en la evolución facial; también, que los humanos presentan altos niveles de reabsorción ósea durante la infancia, una característica que no comparten con otros simios [+].
Una revisión del trabajo de Daver et al. (2022) (que proponía a Sahelanthropus tchadensis como un bípedo habitual) cuestiona tal caracterización, basándose en que determinados rasgos morfológicos, principalmente del fémur, no son exclusivos de seres bípedos [+].
La competencia fue fundamental para la especiación en nuestro camino evolutivo a lo largo de cinco millones de años. Mediante modelos bayesianos y análisis filogenéticos, un trabajo observa que, cuantas más especies de Homo había, mayor era la tasa de especiación. Esto apenas tiene parangón en la ciencia evolutiva dado que, en otros vertebrados, las tasas de especiación aumentan y luego se estancan a medida que los nichos ecológicos se llenan, momento en que las tasas de extinción comienzan a aumentar. Este argumento reforzaría que la competencia entre especies fue un factor evolutivo importante. En cambio, en los homininos, cuando esos nichos se fueron llenando, surgían aún más especies. También lo han observado en algunas especies de escarabajos isleños, cuyos ecosistemas han podido producir tendencias evolutivas inusuales. Los autores de este trabajo observan que, mientras que especies de homininos como Paranthropus probablemente evolucionaron fisiológicamente para ampliar su nicho, por ejemplo, adaptando los dientes para explotar nuevos tipos de alimentos, el motor del patrón tan diferente en nuestro género Homo podría haber sido la tecnología. Las herramientas de piedra, el fuego, o las técnicas de caza intensiva son comportamientos extremadamente flexibles que permiten generalizar y superar rápidamente los nichos ecológicos, que obligan a otras especies a competir por el hábitat y sus recursos, sin necesidad de sobrevivir mucho tiempo para desarrollar nuevos planes corporales [+].
Siempre es bienvenido un nuevo yacimiento olduvayense. Se trata de Sare-Abururu, situado en la península de Homa, en el suroeste de Kenia, donde ya se conocen otros conjuntos olduvayenses procedentes de Nyayanga (Plioceno final, ~3-2,6 Ma) y Kanjera Sur (Pleistoceno inicial, ~2 Ma). Mientras que en estos últimos sitios los homininos obtenían diversas materias primas de alta calidad a más de 10 km de distancia, más tarde, en Sare-Abururu los homininos hace ~1,7 Ma dejaron 1754 artefactos fabricados a partir de guijarros de cuarzo, en una reducción simple aunque produciendo hábilmente bordes cortantes con pocos errores. Esto muestra que las estrategias oportunas no necesariamente indican una falta de habilidad o de materias primas adecuadas. Sare-Abururu se halla a unos 12 km al sureste de Kanjera South y 18 km al este de Nyayanga. Los patrones de adquisición y reducción de líticos en la península de Homa parecen reflejar variaciones en los contextos de recursos locales en lugar de cambios evolutivos a gran escala en la movilidad, el coste energético o la cognición de los fabricantes de herramientas [+].
Herramientas olduvayenses de Sare-Abururu (c. 1.7 Ma). Crédito: Finestone et al. (2024).
Un nuevo trabajo de modelización indica que el Olduvayense pudo haberse originado hace 3-3,2 Ma en algún momento cercano a la ocurrencia de los artefactos de Lomekwi 3 [+].
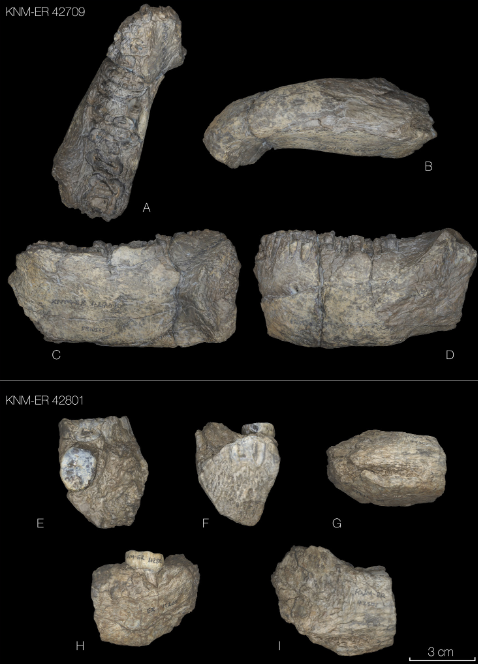
Dos nuevas mandíbulas de Paranthropus boisei aportan información muy interesante sobre esta especie [+]. Por un lado, KNM ER-42709 es una hemimandíbula sin dientes datada en 1,4 Ma, el parántropo más reciente conocido en Kenia hasta ahora, y uno de los más recientes del registro fósil junto con KGA 10-525 de Konso (1,42 Ma) y OH 80 (1,338 Ma). Por otro lado, KNM ER-42801 es un fragmento mandibular con un molar M3 datado en 1,55 Ma. Su entorno se caracterizaba por la abundancia de vegetales tipo C4, en un momento previo al incremento de la cobertura forestal (plantas C3) que tuvo lugar en la transición del Pleistoceno inferior al medio (1,3-0,7 Ma en el este de África). Estos dos especímenes enriquecen la variabilidad intraespecífica en las dimensiones dentales y del cuerpo mandibular.
Dos nuevos fósiles de Paranthropus boisei. Crédito: Lucía Nadal et al (2024)
Se han hallado marcas de corte sobre los restos esqueléticos del elefante FN3-5-MPS (Mammuthus meridionalis), que murió de forma natural, halllado en Fuente Nueva 3, Orce (Granada) entre 2001 y 2003. Fueron realizadas por homininos y por felinos, posiblemente tigres dientes de sable, hace 1,2 Ma (millones de años), según se ha observado en la tafonomía y la microestratigrafía. Se trata de la evidencia más antigua conocida de marcas de cortes en huesos de proboscidios en el oeste de Eurasia [+].
De los tres fósiles originales a partir de los que Dubois definió la especie Homo erectus, un nuevo estudio considera que sólo la calota Trinil 2 corresponde al holotipo [+].
Holotipo de Homo erectus publicado por Dubois (1893): A) molar Trinil 1, B) calota Trinil 2, C) fémur Trinil 3.
En este segundo trimestre del año, el ritmo de publicación de novedades en paleoantropología ha sido realmente alto. Rescato 45 de ellas para elaborar este resumen:
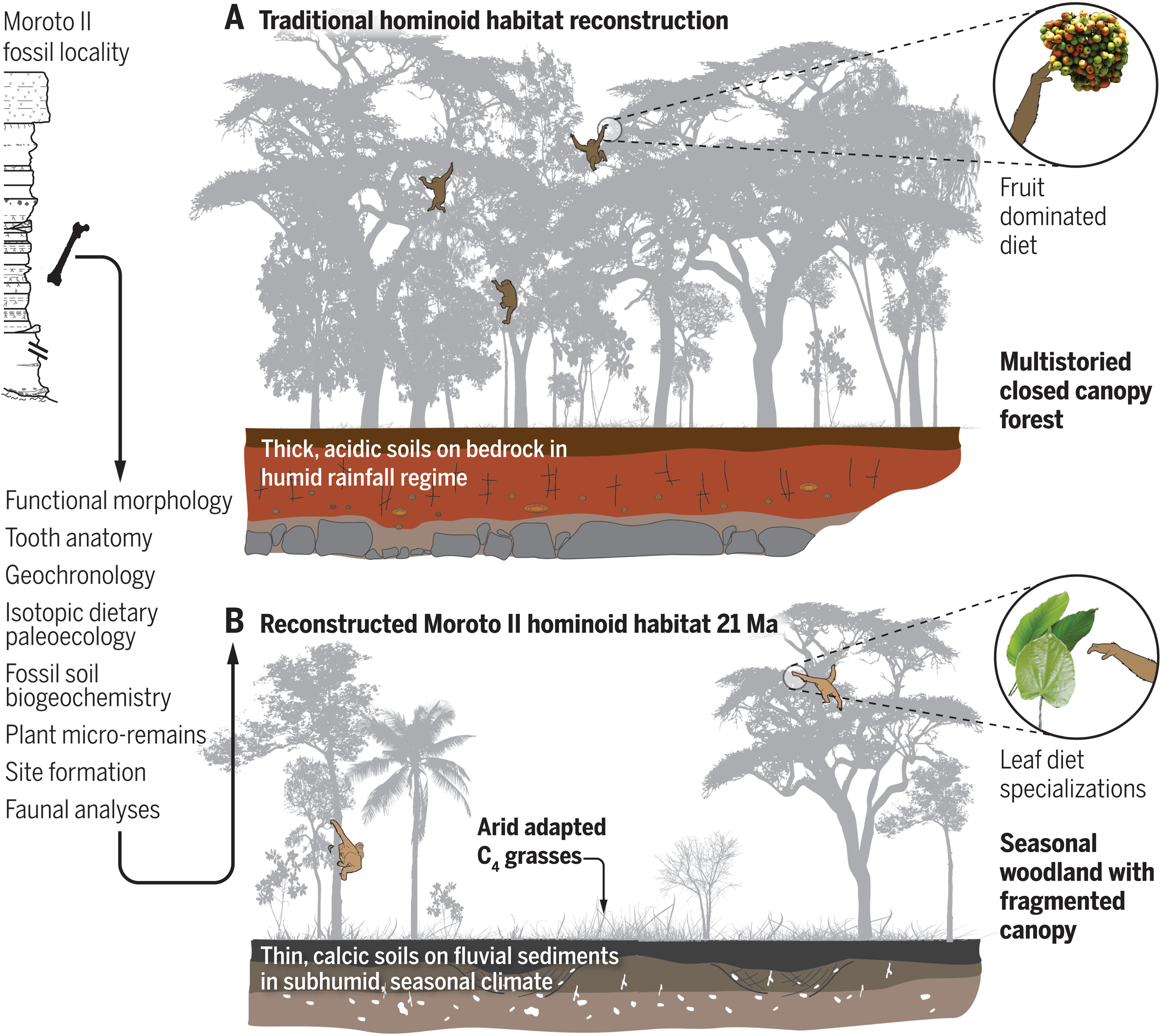
Dos nuevos trabajos paleontológicos y paleoecológicos en el este africano, analizan cómo era el hábitat donde evolucionaron los hominoideos ancestros de los primeros homininos. Aunque se suponía que su ecosistema era predominantente selvático hace más de 10 millones de años (Ma), se ha encontrado que más bien sería una sabana con zonas boscosas, un entorno más seco e irregular. Así, parece que tenemos que buscar el origen de la sabana africana mucho antes antes de lo esperado, con una abundancia de hierbas tipo C4 hace entre 21 y 16 Ma [+]. Esto reabre el debate sobre la evolución morfológica de los hominoideos y el aumento muscular de sus extremidades traseras, observada en el simio más antiguo de la región, Morotopithecus (nombre tomado del sitio Moroto II de Uganda), similar a la de los simios actuales, lo que sería una adaptación a bosques abiertos y estacionales en una transición hacia los orígenes de la locomoción bípeda [+].
Reconstrucción tradicional y nueva del hábitat de los hominoideos. Crédito: L. M. MacLatchy et al (2023).
Un estudio sobre el cúbito de 17 especímenes de homininos (Sahelanthropus, Ardipithecus, Australopithecus, Paranthropus y Homo primitivos), junto con cúbitos de Homo sapiens y de simios extinguidos, concluye que los fósiles TM 266 (asignado a Sahelanthropus tchadensis) y OH 36 (a Paranthropus boisei) se diferencian de otros homininos por encontrarse dentro del morfoespacio de la marcha con nudilleo (knucle-walking). Los autores del trabajo se cuestionan que S. tchadensis fuera un bípedo obligado; más bien podría ser un homínido del Mioceno tardío con adaptaciones para caminar con nudilleo [+].
Se ha logrado crear un modelo digital de la estructura muscular de la parte inferior del cuerpo del esqueleto de Lucy (Australopithecus afarensis), de 3,2 Ma, para comprobar su locomoción en vida. Para ello, se emplearon resonancias magnéticas y tomografías computarizadas de las estructuras musculares y óseas de humanos modernos para trazar las trayectorias musculares, usaron los modelos virtuales del esqueleto de Lucy para recomponer las articulaciones, reconstruyeron 36 músculos, y recrearon sus movimientos en vida. La mayoría de los músculos eran mucho más voluminosos en Lucy que en los humanos modernos. Por ejemplo, los músculos del muslo constituían el 74% de su masa, frente al 50% en los humanos modernos. Los músculos extensores de la rodilla de Lucy, y el efecto de palanca que permitirían, confirman que tenía la misma capacidad de enderezar las articulaciones de la rodilla que los humanos modernos. Una vez más, vemos que Lucy era bípeda [+].
Y, precisamente, los tres Australopithecus afarensis que dejaron las huellas del sitio G de Laetoli hace 3,66 Ma, eran relativamente grandes e iban un poco más rápido de lo que se había estimado anteriormente. Un gran australopiteco (160-170 cm) iba seguido a la misma velocidad por dos individuos de menor tamaño (un posible macho de 140 cm y una hembra o juvenil de 120 cm) [+].
Una revisión sobre la locomoción en los homininos y su evolución desde el Mioceno superior hasta el Plioceno superior, muestra la retención de ciertos rasgos en hombros y extremidades en Au. afarensis y Au. africanus (ya después de ser bípedos habituales) relacionados con actividades arbóreas que resultaban clave para alimentarse, criar y evitar depredadores. Muchos de estos rasgos persistieron hasta la aparición de Homo erectus [+].
Dejamos atrás el tercer trimestre de 2022, con una recopilación de 18 novedades publicadas en este periodo:
Basándose en una veintena de características morfológicas y biomecánicas estudiadas, Sahelanthropus tchadensis parece confirmarse como un bípedo habitual hace unos 7 millones de años (Ma) cuando caminaba sobre el suelo y también sobre las ramas de los árboles, mientras que los huesos del antebrazo indican que era cuadrúpedo para trepar a los árboles, pero de manera distinta al chimpancé y al gorila [+info].
Restos poscraneales de Sahelanthropus tchadensis: fémur TM 266-01-063, y cúbitos TM 266-01-358 y TM 266-01-050 / crédito: Guillaume Daver et al, 2022. Cráneo TM 266-01-060-1 / crédito: Roberto Sáez.
De acuerdo a un estudio experimental, las primeras técnicas para fabricar herramientas de piedra hace 2,6 Ma pudieron reinventarse sin necesidad de una transmisión cultural. Esta pudo haber comenzado mucho más tarde [+info].
La revisión de 23 dientes poscaninos de diferentes cuevas sudafricanas, atribuidos a Homo tempranos, muestra que solo 4 de ellos pueden ser atribuidos sin ambigüedad (3 de Swartkrans y 1 de Sterkfontein), mientras que otros 3 de Sterkfontein se aproximan más a Homo y el resto tienen más rasgos australopitecinos [+info].
Un modesto molar encontrado en el nuevo sitio de Qvemo Orozmani, a unos 20 km de Dmanisi (Georgia) constituye el resto humano más antiguo de Eurasia fuera de Dmanisi, ya que se le estima una datación de similar de 1,77-1,84 Ma a los restos de este sitio [+info].
Orozmani: molar encontrado en 2022 (foto: REUTERS/David Chkhikvishvili).
Hace casi dos años, nos quedamos con la noticia de que Sahelanthropus tchadensis podría en realidad no ser bípedo y por tanto no ser un hominino (Macchiarelli et al, 2020). Tradicionalmente, en los textos sobre evolución humana se ha incluido a esta especie como el representante más antiguo conocido de los homininos (perteneciente a la filogenia humana, que se separó del linaje de los chimpancés hace unos 6-7 Ma, millones de años), principalmente por el foramen magnum (orificio donde se inserta la columna vertebral) en una posición más centrada en el cráneo que los primates cuadrúpedos (Brunet, 2002), lo que corresponde a un ser bípedo.
La bipedación es un factor clave para que una especie se considere como hominina, junto con la reducción del canino y la pérdida del complejo canino-premolar. Precisamente, la datación estimada para los fósiles de Sahelanthropus tchadensis coincide aproximadamente con esa cronología; de ahí la importancia de esta especie para entender nuestra evolución, y para estar estudiando una rama de nuestro árbol muy cercana al Último Ancestro Común de humanos y chimpancés. Tan solo otra especie se aproxima a dicha cronología, Orrorin tugenensis, cuyos restos de fémur sugieren que era un ser bípedo, pero hasta 2020 no se había publicado el fémur de Sahelanthropus, elemento esquelético fundamental para entender su tipo de locomoción. Por la obvia limitación del registro fósil del Mioceno, la dificultad de clasificar unos fósiles dentro o fuera del linaje hominino es enorme, y las dudas sobre su bipedación la empeoran.
Un fémur parcial y dos cúbitos se hallaron en 2001 en Toros-Menalla, Chad, cerca del lugar donde se recuperó el holotipo de Sahelanthropus tchadensis en el mismo año, el icónico cráneo apodado Toumaï («esperanza de vida» en dazaga, lengua local de Chad). También se encontraron algunos dientes y fragmentos mandibulares. Aunque no se puede asegurar que los restos poscraneales y el cráneo sean del mismo individuo, sí que hay un consenso alrededor de que pertenecen a la misma especie S. tchadensis. Además, las diferencias respecto al fémur de Orrorin tugenensis (BAR 1002’00) garantizan que efectivamente ambos huesos son de especies diferentes. Sin embargo, Macchiarelli y otros planteaban serias dudas respecto a que los huesos poscraneales fuera de un individuo bípedo habitual, sino más bien de un grupo derivado de Pan (chimpancés), o de una radiación adaptativa de homínidos africanos acaecida entre el Mioceno final (que termina hace 5,3 Ma) y el Plioceno inicial, con grupos extinguidos que tendrían una morfología novedosa y una mezcla de rasgos primitivos y derivados que recuerdan en parte a homininos y a paninos.
Restos poscraneales de Sahelanthropus tchadensis: fémur TM 266-01-063, y cúbitos TM 266-01-358 y TM 266-01-050. Crédito: Guillaume Daver et al, 2022.
Poco después de aquello, Franck Guy, Guillaume Daver, y otros publicaron un preprint donde analizaban los materiales originales del mismo fémur izquierdo y de dos cúbitos, en comparación con otros primates (chimpancé, gorila, orangután, simios del Mioceno, Orrorin, Ardipithecus, Australopitecus, especies extintas de humanos y Homo sapiens). Este trabajo es el ahora publicado en Nature (Daver, Guy, Mackaye et al, 2022), y que ha despertado tanta atención entre la comunidad científica y el público general. Basándose en una veintena de características morfológicas y biomecánicas estudiadas, Daver y otros sostienen que S. tchadensis era efectivamente un bípedo habitual cuando caminaba sobre el suelo y también sobre las ramas de los árboles, mientras que los huesos del antebrazo indican que era cuadrúpedo para trepar a los árboles, pero de manera distinta al chimpancé y al gorila.
Un trabajo posterior (Williams et al., 2026) analiza los fragmentos de cúbitos y fémur de Toumaï mediante morfometría geométrica 3D y rasgos cualitativos. Aunque la curvatura de los ejes y robustez de estos huesos se asemejan más al chimpancé entre primates vivos y fósiles, la propoción ulna-fémur es intermedia entre bonobos y Australopithecus (StW 573). Además, tres rasgos femorales se alinean con adaptaciones bipedales propias de los homininos: 1) un tubérculo femoral prominente para la inserción del ligamento iliofemoral, que estabiliza la cadera extendida cuando el tronco está erguido sobre piernas extendidas; 2) una fuerte antetorsión diafisaria, correlacionada con un ángulo bicondilar valgo para alinear rodilla y tobillo bajo el centro de masa; 3) un complejo glúteo derivado, con tuberosidad glútea y cresta sin línea espiral lateral, similar a Orrorin tugenensis y Ardipithecus ramidus, que ordena la musculatura de cadera para la postura erguida.