
En este segundo trimestre del año, el ritmo de publicación de novedades en paleoantropología ha sido realmente alto. Rescato 45 de ellas para elaborar este resumen:
- Dos nuevos trabajos paleontológicos y paleoecológicos en el este africano, analizan cómo era el hábitat donde evolucionaron los hominoideos ancestros de los primeros homininos. Aunque se suponía que su ecosistema era predominantente selvático hace más de 10 millones de años (Ma), se ha encontrado que más bien sería una sabana con zonas boscosas, un entorno más seco e irregular. Así, parece que tenemos que buscar el origen de la sabana africana mucho antes antes de lo esperado, con una abundancia de hierbas tipo C4 hace entre 21 y 16 Ma [+]. Esto reabre el debate sobre la evolución morfológica de los hominoideos y el aumento muscular de sus extremidades traseras, observada en el simio más antiguo de la región, Morotopithecus (nombre tomado del sitio Moroto II de Uganda), similar a la de los simios actuales, lo que sería una adaptación a bosques abiertos y estacionales en una transición hacia los orígenes de la locomoción bípeda [+].
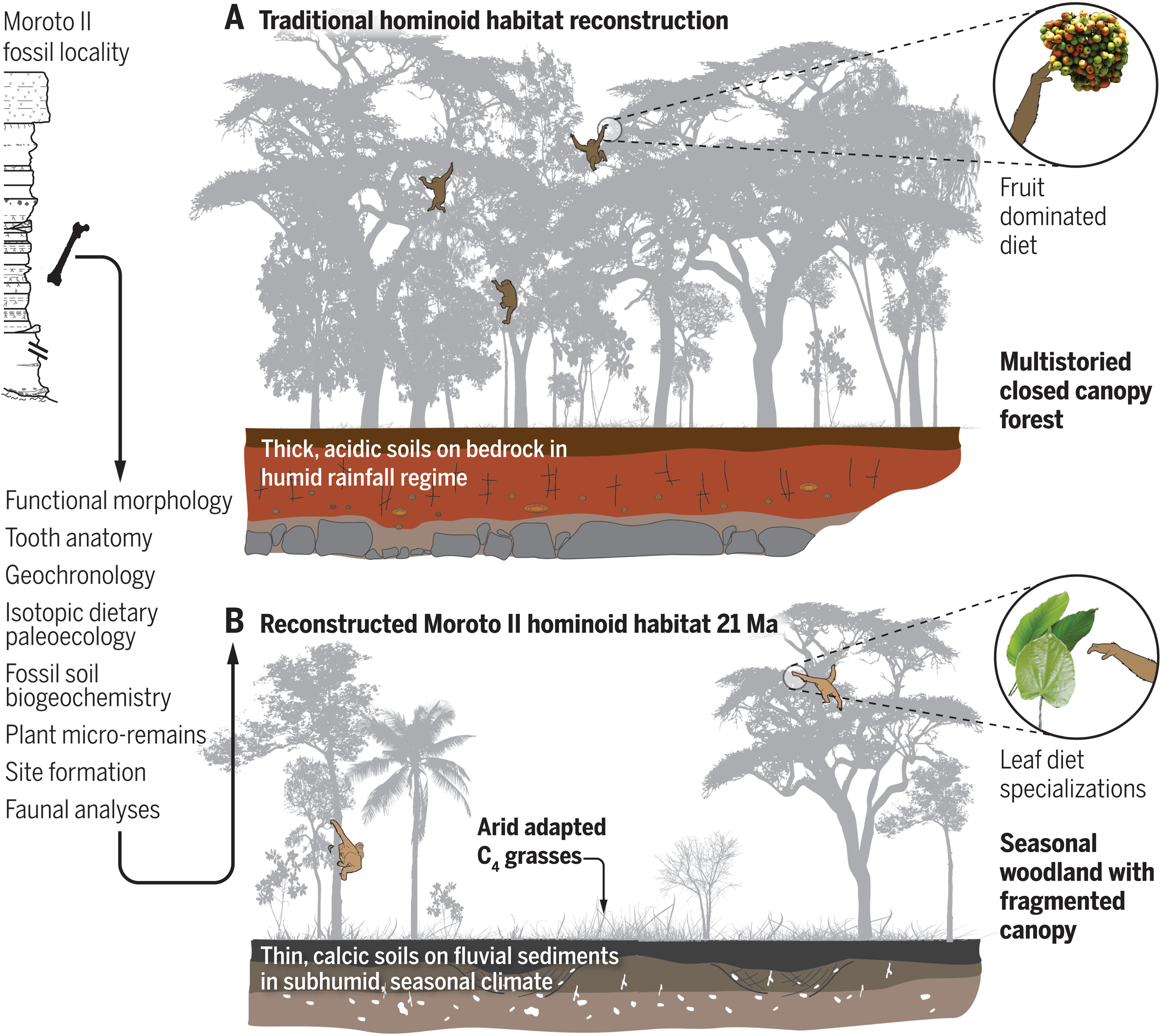
Reconstrucción tradicional y nueva del hábitat de los hominoideos. Crédito: L. M. MacLatchy et al (2023).
- Un estudio sobre el cúbito de 17 especímenes de homininos (Sahelanthropus, Ardipithecus, Australopithecus, Paranthropus y Homo primitivos), junto con cúbitos de Homo sapiens y de simios extinguidos, concluye que los fósiles TM 266 (asignado a Sahelanthropus tchadensis) y OH 36 (a Paranthropus boisei) se diferencian de otros homininos por encontrarse dentro del morfoespacio de la marcha con nudilleo (knucle-walking). Los autores del trabajo se cuestionan que S. tchadensis fuera un bípedo obligado; más bien podría ser un homínido del Mioceno tardío con adaptaciones para caminar con nudilleo [+].
- Se ha logrado crear un modelo digital de la estructura muscular de la parte inferior del cuerpo del esqueleto de Lucy (Australopithecus afarensis), de 3,2 Ma, para comprobar su locomoción en vida. Para ello, se emplearon resonancias magnéticas y tomografías computarizadas de las estructuras musculares y óseas de humanos modernos para trazar las trayectorias musculares, usaron los modelos virtuales del esqueleto de Lucy para recomponer las articulaciones, reconstruyeron 36 músculos, y recrearon sus movimientos en vida. La mayoría de los músculos eran mucho más voluminosos en Lucy que en los humanos modernos. Por ejemplo, los músculos del muslo constituían el 74% de su masa, frente al 50% en los humanos modernos. Los músculos extensores de la rodilla de Lucy, y el efecto de palanca que permitirían, confirman que tenía la misma capacidad de enderezar las articulaciones de la rodilla que los humanos modernos. Una vez más, vemos que Lucy era bípeda [+].
- Y, precisamente, los tres Australopithecus afarensis que dejaron las huellas del sitio G de Laetoli hace 3,66 Ma, eran relativamente grandes e iban un poco más rápido de lo que se había estimado anteriormente. Un gran australopiteco (160-170 cm) iba seguido a la misma velocidad por dos individuos de menor tamaño (un posible macho de 140 cm y una hembra o juvenil de 120 cm) [+].
- Una revisión sobre la locomoción en los homininos y su evolución desde el Mioceno superior hasta el Plioceno superior, muestra la retención de ciertos rasgos en hombros y extremidades en Au. afarensis y Au. africanus (ya después de ser bípedos habituales) relacionados con actividades arbóreas que resultaban clave para alimentarse, criar y evitar depredadores. Muchos de estos rasgos persistieron hasta la aparición de Homo erectus [+].
- Se han analizado 124 huesos fósiles de la cueva de Drimolen (Sudáfrica) datados en 1,95-2,04 Ma, comparándolos con estudios tafonómicos, experimentales y datos en la literatura tafonómica. El trabajo identifica 51 de ellos como herramientas óseas, con una punta redondeada y un patrón de uso característicos de su empleo por homininos. Posiblemente se emplearon para múltiples aplicaciones. Así, se amplía a 65 el registro de herramientas óseas conocidas en el lugar [+].
- Seguimos en las cuevas sudafricanas. Se han estudiado cuatro individuos infantiles de Paranthropus robustus, tres de la unidad P de Kromdraai (KW 6420, KW 9000/9600 y KW 10840) y uno de Drimolen (DNH 47). Los hallazgos son: 1) aunque la mayoría de los rasgos craneofaciales robustos distintivos aparecen relativamente tarde en la ontogenia, unos pocos no lo hacen; 2) inesperadamente, el crecimiento de las regiones premaxilar y maxilar tienen un patrón independiente; 3) el crecimiento diferencial da lugar a una fosa cerebral proporcionalmente más grande y más rotada posteroinferiormente en los individuos infantiles de P. robustus que en el Niño de Taung (Australopithecus africanus); 4) P. robustus parece más estrechamente relacionado con Homo que con A. africanus; 5) es probable que la icónica calota juvenil de SK 54 (al que se comió un leopardo) fuera de Homo, en vez de Paranthropus [+]

4 individuos infantiles de Paranthropus robustus. Crédito: J. Braga et al (2023).
- Sobre el debate de la participación de la mujer en la caza durante la prehistoria, un trabajo sobre 63 sociedades con economía de forrajeo de los últimos 100 años analiza dicha participación, como posible referencia proyectable al pasado. 19 de las sociedades analizadas son de Norteamérica, 6 de Sudamérica, 12 de África, 15 de Australia, 5 de Asia y 6 del resto de Oceanía. En 36 de ellas se documenta caza intencional por parte de mujeres, en otras 5 la caza es oportunista, en 9 no hay datos sobre si es intencional u oportunista, y en 13 no hay datos de que las mujeres cacen. De 45 sociedades con datos del tamaño de piezas cazadas, en 21 de ellas las mujeres cazan piezas de tamaño pequeño, en 7 de tamaño mediano, en 15 de tamaño grande, y en 2 variable [+]. Son resultados que vienen a mostrar la variabilidad de modos económicos en distintas regiones geográficas, y en los que en general hay una participación de las mujeres en la caza (en 50 de 63 sociedades), mayoritariamente de piezas pequeñas (en 21 de 45) pero también una proporción notable de piezas grandes (en 15 de 45).
- A partir de datos empíricos, se ha comprobado que el cabello muy rizado proporciona la protección más eficaz para el cuero cabelludo frente a la radiación solar, al tiempo que minimiza la necesidad de sudor para compensar la ganancia de calor. La postura bípeda y un cuerpo sin pelo pudieron haber hecho necesario el desarrollo de pelo en el cuero cabelludo para minimizar la ganancia de calor de la radiación solar, particularmente en homininos con grandes cerebros [+].
- ¿Un caso muy antiguo de antropofagia? En la tibia de hominino KNM-ER 741 (de Homo habilis o Paranthropus boisei, sin claridad sobre su asignación a una especie), existen 11 marcas de las cuales se han identificado 9 como marcas de corte y 2 como marcas de dientes. Los dientes parecen de un león que consumió la masa muscular principal, y luego los homininos carroñaron los restos de carne del extremo de la tibia. Esto ocurrió hace 1,45 Ma en Koobi Fora, Kenia [+].

Tibia ER 741 con marcas de corte. Crédito: Pobiner, B., Pante, M. y Keevil, T. (2023)
- Mientras que la aparición del Achelense está bien documentada en África Oriental en ~1,7 Ma, los desarrollos posteriores son menos conocidos y hasta cierto punto controvertidos. Se ha conseguido estudiar la evolución de la tecnología en el barranco Gombore de Melka Kunture (Etiopía) en el periodo 1,6-1,2 Ma, donde el paleoambiente difirió significativamente del de otros lugares de África (por su elevación a 2000 m, el clima más fresco y lluvioso, el desarrollo de especies animales y vegetales endémicas y el impacto recurrente de la actividad volcánica). En Gombore IB (1,6 Ma) hay restos de Homo erectus/ergaster asociados a un rico conjunto achelense temprano. Pero en Gombore Iγ (1,4 Ma) y Gombore Iδ (1,3 Ma), en contraste con el sondeo C de Gombore OAM (1,2 Ma), hay escasez o ausencia de grandes lascas y large cutting tools, posiblemente como adaptación de H. erectus/ergaster a los recursos locales, en un entorno relativamente aislado [+].
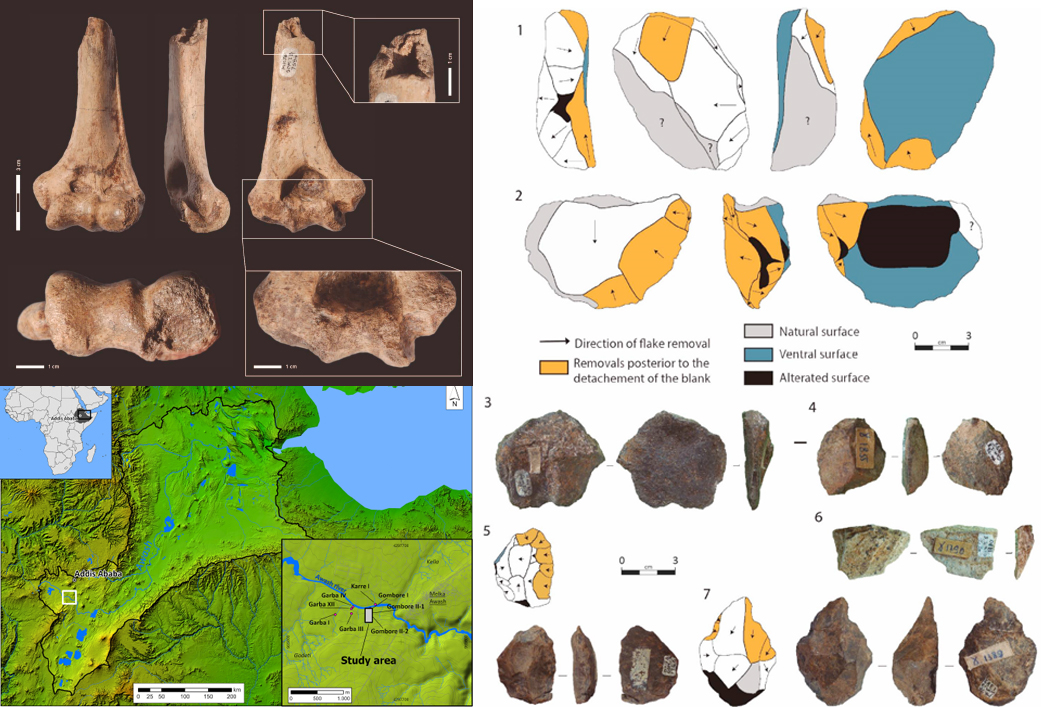
Sitios y materiales de Gombore. Crédito: M. Mussi et al (2023)
- Mientras tanto en Europa, continúan los trabajos en localidades al aire libre en el entorno del Barranc de la Boella (Tarragona, España), que tienen el registro achelense más antiguo conocido en Europa (0,99-0,78 Ma). Los resultados revelan un conjunto tecnológico diferente del resto de yacimientos de este continente, que son del Pleistoceno medio. Esto respalda la hipótesis de que el Barranc de la Boella puede representar una dispersión achelense temprana, no reconocida hasta ahora, que salió de África conectada con sus primeras evidencias a las puertas de Eurasia, y se desplazó por la ruta costera del norte del Mediterráneo para llegar a Europa occidental [+].
- Se ha anunciado la evidencia de presencia humana más antigua de Grecia: un conjunto de herramientas del Paleolítico inferior en el yacimiento Kyparissia 4 (Megápolis, Arcadia) con una antigüedad inicialmente estimada de 700 ka, aunque pendientes de datación y de su publicación [+].
- Los bifaces y hendidores fueron un desarrollo continuo en Europa occidental a lo largo del tiempo por acciones acumulativas, con estrategias largas y complejas, y no tanto por innovaciones repentinas, mostrando dos grandes grupos tecnológicos: el noroeste y centro de Francia y Gran Bretaña (MIS 17/16 a MIS 11) y el borde atlántico (Atapuerca en España y Menez-Dregan en Francia, desde MIS 12/11 a MIS 8) [+].
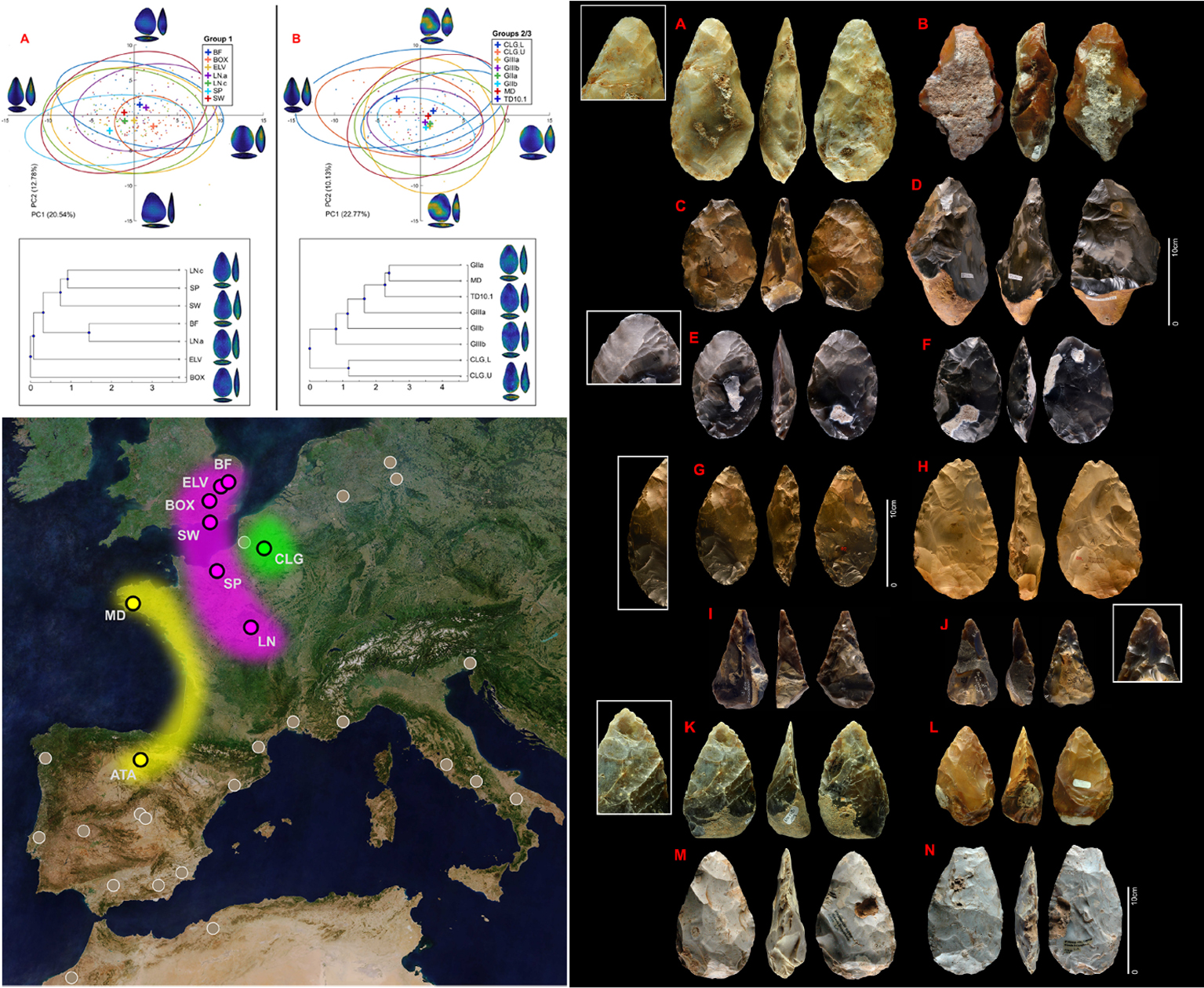
Estudio sobre el Achelense de Europa occidental. Crédito: Paula García-Medrano et al (2023)
- Un nuevo estudio de las mandíbulas de la Sima de los Huesos (Atapuerca), las acercan (todavía más) al linaje neandertal, y las alejan (por si quedaban dudas) del holotipo de Homo heidelbergensis, la mandíbula de Mauer [+].
- Y la evaluación de las vértebras cervicales de la Sima de los Huesos (172 especímenes, pertenecientes a un mínimo de 11 atlas, 13 axis y 52 subaxiales) muestra más similitudes con el linaje neandertal que con el sapiens, pero también diferencias respecto a la morfología neandertal, principalmente en cuanto a longitud y robustez en las vértebras cervicales subaxiales inferiores, posiblemente relacionadas con el aumento del tamaño del cerebro y/o cambios en la morfología craneal que se produjeron en el linaje neandertal [+].
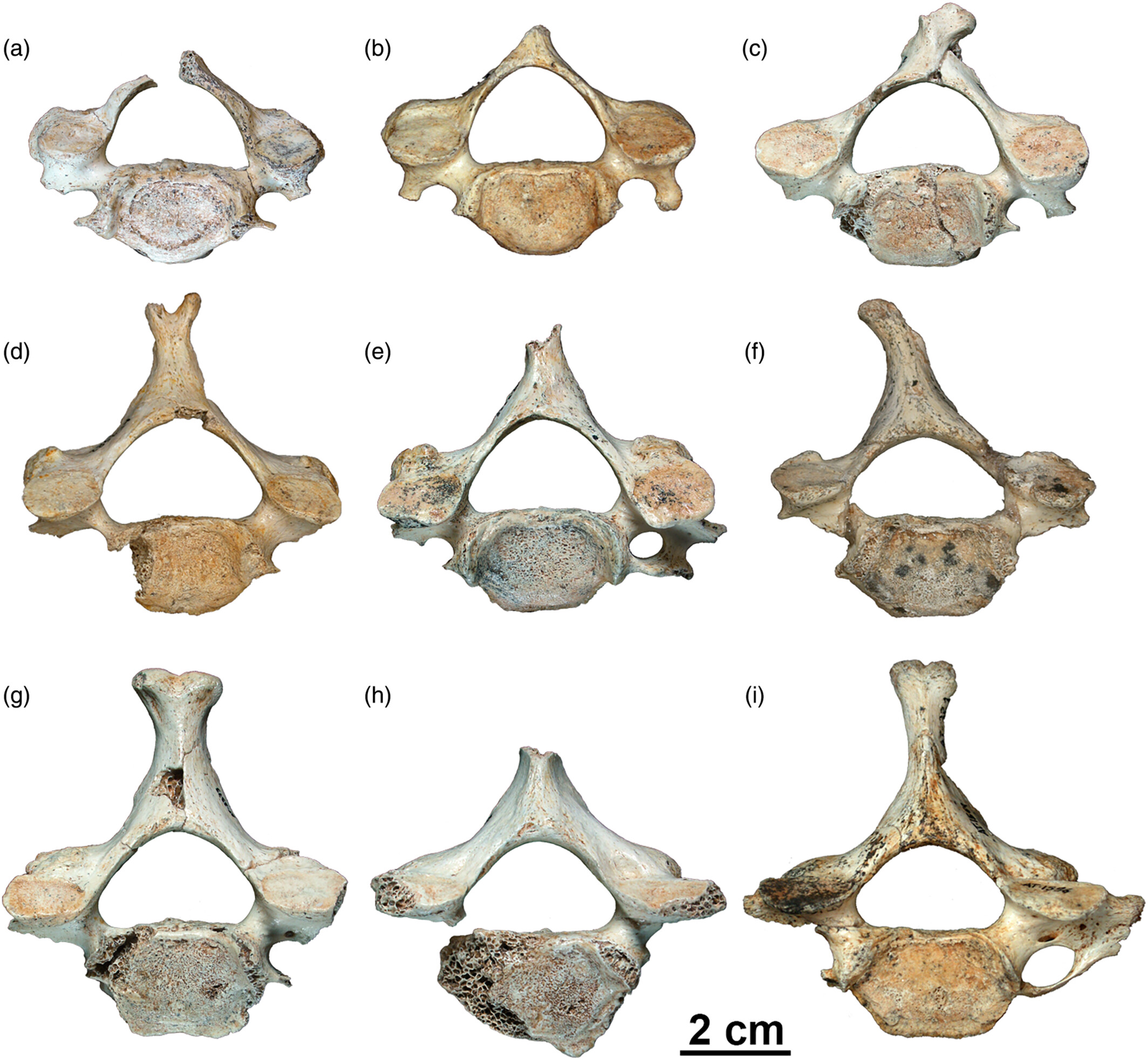
Vértebras cerviales subaxiales más completas en el registro de la Sima de los Huesos. Crédito: Gómez-Olivencia y Arsuaga (2023)
- Hace 300 ka en Schöningen (Alemania), manadas de elefantes, rinocerontes y otras especies de herbívoros se congregaron a lo largo de un paleolago junto a un bosque rico en abedules, pinos y gramíneas, dejando sus huellas en dos sitios, Schöningen 13 I-Fs2 y Schöningen 13 II-2 Untere Berme. En este último yacimiento se han registrado además tres posibles huellas de homininos, las más antiguas conocidas en Alemania [+].

Huellas fósiles (y una de las posibles huellas humanas) de Schöningen 13 II-2 Untere Berme. Crédito: Senckenberg
- Un trabajo de modelización de posibles escenarios de origen y evolución de Homo sapiens en África, indica que el más probable es el desarrollo de dos linajes troncales poco estructurados (hace al menos 400 ka) que se mantuvieron en contacto genético durante cientos de milenios. Estos modelos explicarían patrones de polimorfismo que anteriormente se habían atribuido a contribuciones de homininos arcaicos en África. Hace 120-135 ka se produjo una divergencia entre esas dos poblaciones: una dio lugar a grupos en el sur, este y oeste africanos, y la otra se extinguió, tras mezclarse con algunas de los grupos descendientes de la primera, sobre todo en el oeste. Un 1-4% de la diferenciación genética entre las poblaciones humanas contemporáneas puede atribuirse a la deriva genética entre las dos poblaciones madre [+].
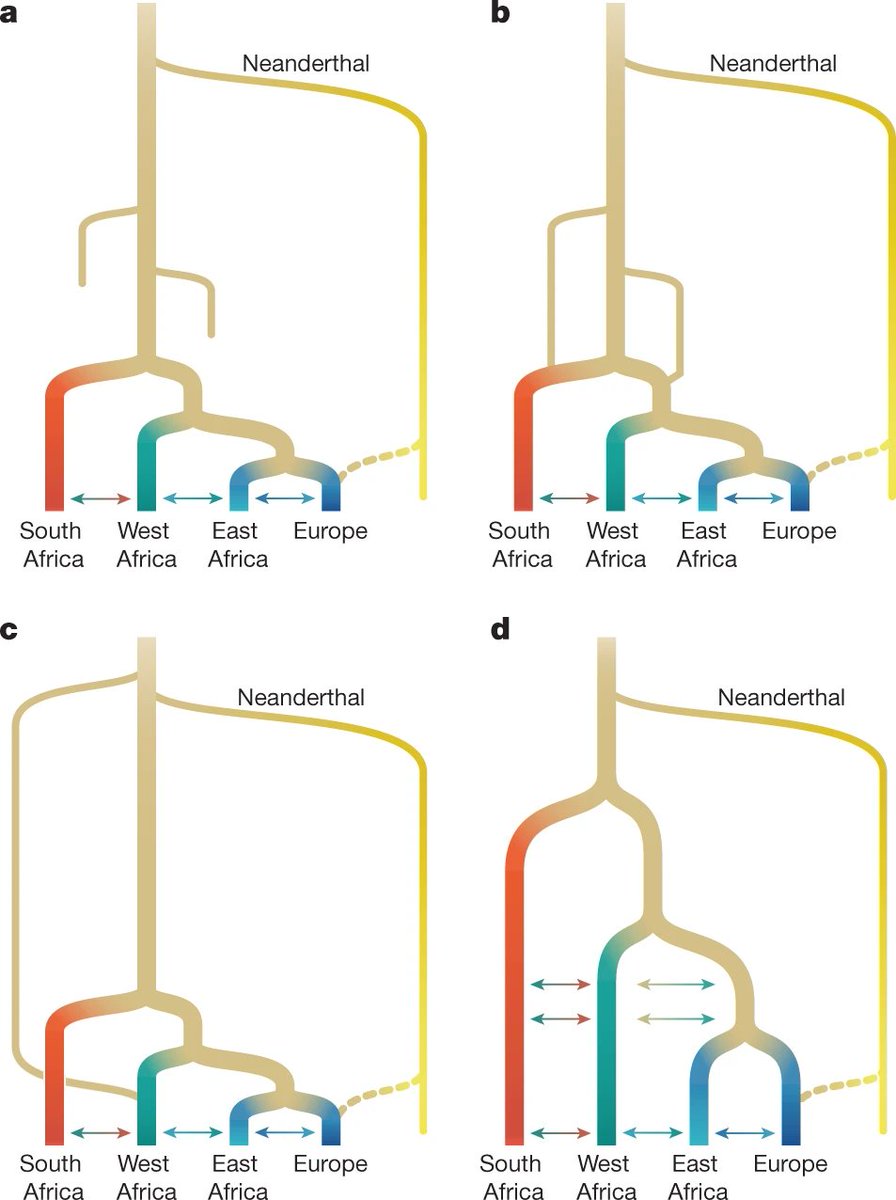
Modelos conceptuales para la evolución de Homo sapiens en África. a-Expansión reciente b-ídem con persistencia regional c-Mezcla arcaica d-Multirregional africano. Crédito: Stancampiano, L.M., Rubio-Jara, S., Panera, J. et al. (2023)
- La tecnología MSA encontrada en Bargny (costa de Senegal, oeste de África) presenta características comunes con las observadas en otros sitios del continente en el Pleistoceno medio tardío, pero que se mantienen estables de manera única en el oeste africano hasta el inicio del Holoceno. Se trata de un sitio húmedo, donde los microfósiles vegetales que indican la proximidad de hábitats estuarinos. La habitabilidad persistente de los entornos de África occidental, incluidos los manglares, contribuye a trayectorias de estabilidad conductual claramente africanas [+].
- Se ha publicado el catálogo descriptivo completo de los 150 dientes de Homo naledi recuperados en las excavaciones de 2013 a 2015 de la cámara Dinaledi, sitio UW 101, dentro del sistema de cuevas Rising Star (Sudáfrica), datados en 330-241 ka [+].
- Un conjunto de tres estudios (pendientes de revisión por pares) plantean capacidades cognitivas avanzadas en Homo naledi plasmadas en la cámara Dinaledi y en la antecámara Hill a través de posibles enterramientos y el trazado de grabados rupestres [+].

(1,2) Dos zonas de enterramiento en la cámara Dinaledi. (3,4) Zona de enterramiento en la antecámara Hill. Crédito: Berger L. et al., 2023. | Rising Star program | National Geographic
- El estudio de subproductos de la combustión en Valdocarros II (España) sugieren la presencia de fuegos antropogénicos controlados hace unos 245 ka, a baja temperatura (alrededor de 350ºC, frente a un fuego no controlado que puede alcanzar los 800ºC, el cual carboniza el exterior de la carne pero el interior queda crudo). Empleaban hidrocarburos poliaromáticos, producto de la combustión incompleta (350ºC) de ramas de pino secas, que no abundaban en su ecosistema, lo que sugiere una búsqueda intencionada. Valdocarros II es uno de los mayores yacimientos achelenses excavados en la península ibérica, junto con Torralba y Ambrona (Soria), y muestra numerosos conjuntos de restos faunísticos y artefactos achelenses en cinco niveles. Este caso uno de los pocos ejemplos registrados de fuego antropogénico del Pleistoceno medio en Europa hasta la fecha, y una de las evidencias más antiguas de uso del fuego en un yacimiento achelense al aire libre en Europa junto con Terra Amata en Francia, y el único en la península ibérica [+].

Restos de fuego de 245 ka en Valdecarros II. Crédito: Stancampiano, L.M., Rubio-Jara, S., Panera, J. et al. (2023)
- Un trabajo sobre la tecnología y las estrategias de movilidad de los grupos neandertales que habitaron la cueva de Ciota Ciara (noroeste de Italia) durante el Pleistoceno medio, ha identificado un conjunto de artefactos «exóticos» (lascas sin retocar, útiles retocados y, esporádicamente, pequeños núcleos) elaborados con materias primas tomadas a entre 2 y 30 km de distancia, e introducidos en el sitio como herramientas móviles para distintas funciones, principalmente para procesar carne [+]. Se trata de un estudio importante para entender los conjuntos de herramientas móviles, y el debate entre herramientas de oportunidad (improvisadas) y herramientas elaboradas o formales (conservadas).
- Con 153 ka, esta huella del Parque Nacional Garden Route (Sudáfrica) es la más antigua atribuida a nuestra especie Homo sapiens [+].

- Mediante el estudio de isótopos en la corona dental, se ha podido determinar la alimentación que tenían los humanos que vivían en sistema de cuevas de Almonda (Portugal) en el Paleolítico medio y en el Magdaleniense. Hace unos 100 ka, los neandertales comían cabras en verano y fauna de mayor tamaño en invierno (caballo, ciervo, rinoceronte), que iban a buscar a largas distancias, abarcando unos 600 km2. Los humanos del Magdaleniense se movían estacionalmente, comían conejos, ciervos, cabras monteses y peces de agua dulce, y abarcaban un territorio más pequeño (unos 300 km2) [+], posiblemente porque el aumento en la densidad de población en el Paleolítico superior pudo reducir el territorio disponible.
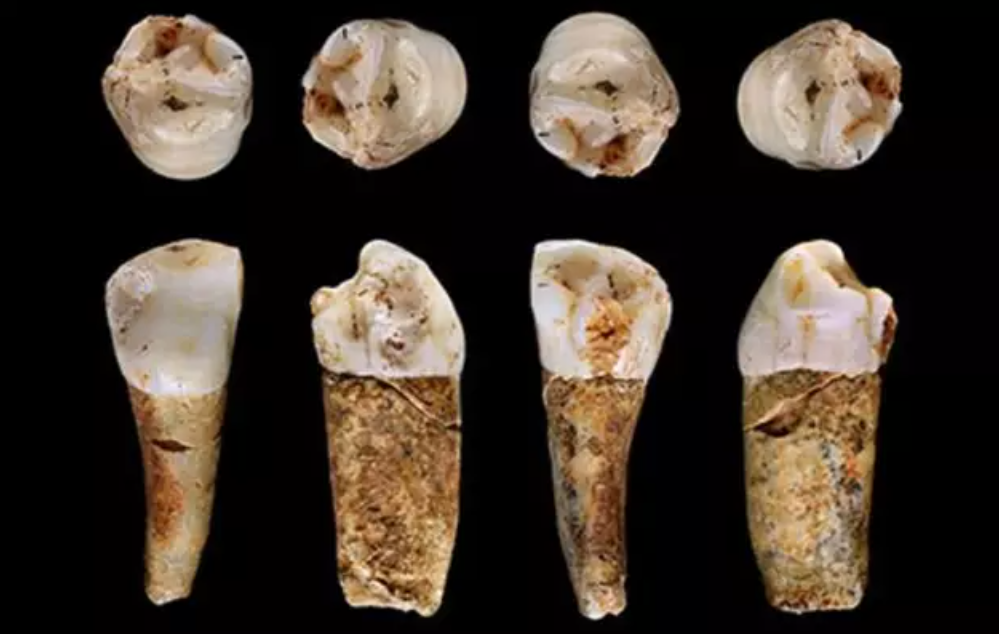
Premolar neandertal de Almonda. Crédito: B. Linscott et al (2023).
- Un nuevo estudio demuestra la complejidad de la producción de brea de abedul por parte de neandertales en Königsaue (Alemania) hace entre 45-80 ka, mediante cocción reductora controlada. Destilaron brea en un ambiente subterráneo creado intencionadamente, que restringió el flujo de oxígeno y permaneció invisible durante el proceso. Es poco probable que este grado de complejidad se inventara espontáneamente, sino que los neandertales inventaron o desarrollaron este proceso basándose en métodos anteriores más simples, un indicador claro de evolución cultural acumulativa en el Paleolítico medio europeo [+].
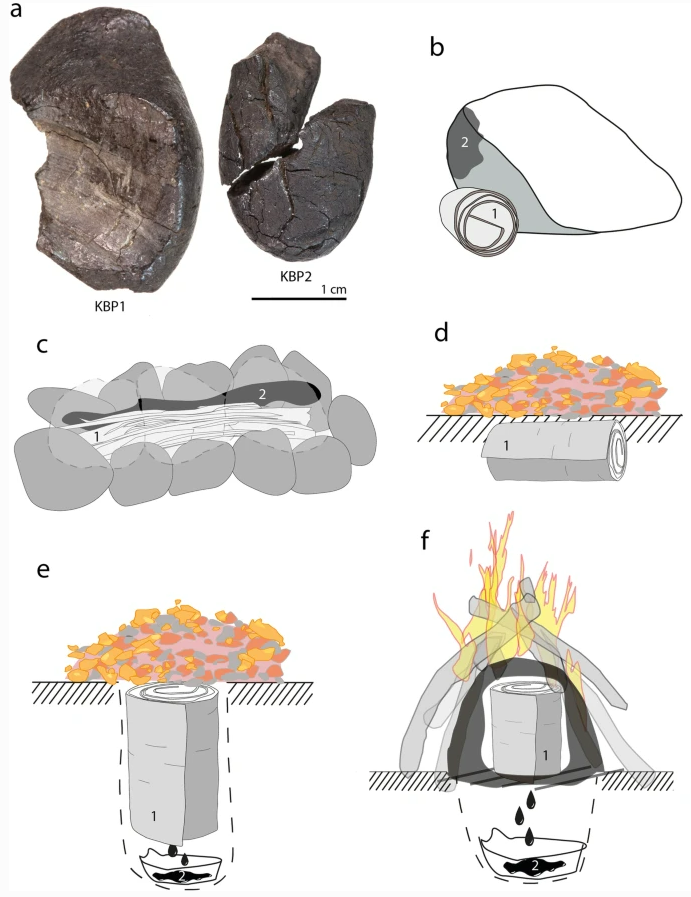
Brea de abedul de Königsaue y técnicas de producción experimentales. Crédito: P. Schmidt et al (2023)
- Se ha observado el contacto entre dos poblaciones culturalmente distintas de neandertales con orígenes diferentes habitaron el norte del Cáucaso (una región desde el MIS 5 hasta el final del MIS 3, momento de desaparición de esta especie [+]. El Paleolítico medio está representado en el Cáucaso entre hace 260-210 ka y 40 ka aproximadamente, con diversas poblaciones de homininos, pero la compleja historia de la población de la región durante este periodo sigue siendo poco conocida.
- Una datación importante: 86-68 ka es la antigüedad de los fragmentos de hueso frontal (TPL 6) y de tibia (TPL 7) de Homo sapiens procedentes de la cueva Tam Pà Ling, en el norte de Laos. La morfología del frontal sugiere la descendencia de una población inmigrante grácil más que la evolución o la mezcla con poblaciones arcaicas locales. Este hallazgo supone un gran aporte para entender la presencia antigua de nuestra especie fuera de África, junto con la mandíbula de Misliya, Israel, de 180 ka, la falange de Al Wusta, Arabia Saudí, de 90 ka, y los restos de los sitios chinos de Fuyan Cave (120-80 ka), Huanglongdong (100-80 ka), Lunadong (127-70 ka) y Zhirendong (116-106 ka), aunque con dudas en algunas de las últimas dataciones [+]. Tal vez recordaréis que el preprint de este trabajo se había publicado en el trimestre anterior.
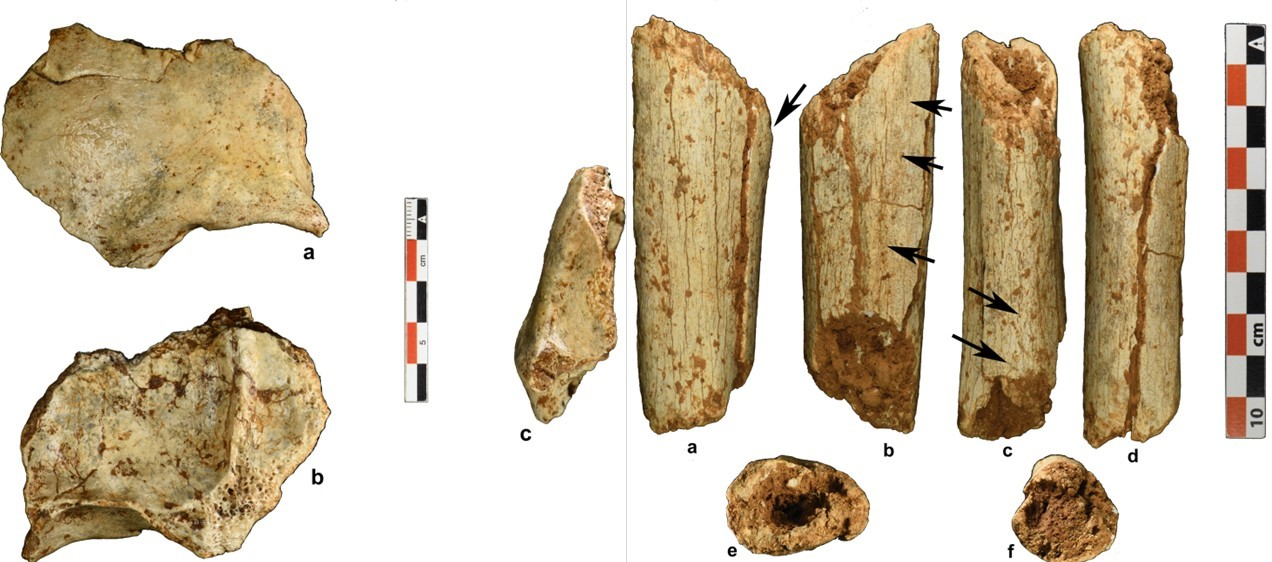
Fragmentos de frontal y de tibia de Homo sapiens procedentes de Tam Pà Ling (norte de Laos). Crédito: Freidline, S.E., Westaway, K.E., Joannes-Boyau, R. et al. (2023).
- Se ha publicado un conjunto de 103 herramientas de hueso realizadas por los grupos neandertales en Chez-Pinaud (Francia) hace 60-65 ka, de las que el 20% tienen evidencias de reutilización. Este hallazgo, sumado al conjunto de 1200 útiles óseos de 50-60 ka procedente de Chagyrskaya (Altai, Siberia, Rusia), estudiado por el mismo equipo, así como otros yacimientos dispersos, sigue ayudando a descartar la asociación exclusiva de la tecnología ósea a nuestra especie que se suponía hasta no hace mucho tiempo [+].

Ejemplo de útil óseo hecho por neandertales en Chez-Pinaud. Crédito: Baumann M, Plisson H, Maury S, Renou S, Coqueugniot H, Vanderesse N, et al. (2023)
- Se ha presentado un conjunto de grabados realizados por neandertales con marcas de dedos hace al menos 57 ka en la cueva de La Roche-Cotard, en el centro del Valle del Loira en Francia. En una pared de 12 metros, hay un gran número de trazos de dos tipos: puntos más o menos circulares y simples hechos con la punta del dedo, que apuntan en la misma dirección, y líneas más complejas dejadas al arrastrar la punta del dedo por la superficie de la pared, que forman distintas estructuras en paneles triangulares y rectangulares [+].

Uno de los paneles de grabados neandertales en La Roche-Cotard. Crédito: J-C Marquet et al (2023)
- Se han hallado los restos de un cuarto individuo neandertal en Cova de les Teixoneres, Moià (España). Tiene unos 52 ka. Son dos fragmentos de occipital de un juvenil, que se unen a los otros individuos que comenzaron a aparecer en 2016: otro niño, un adulto joven y un adulto. Se han identificado marcas de corte sobre la superficie de algunos huesos que se interpretan como actividades de canibalismo [+].
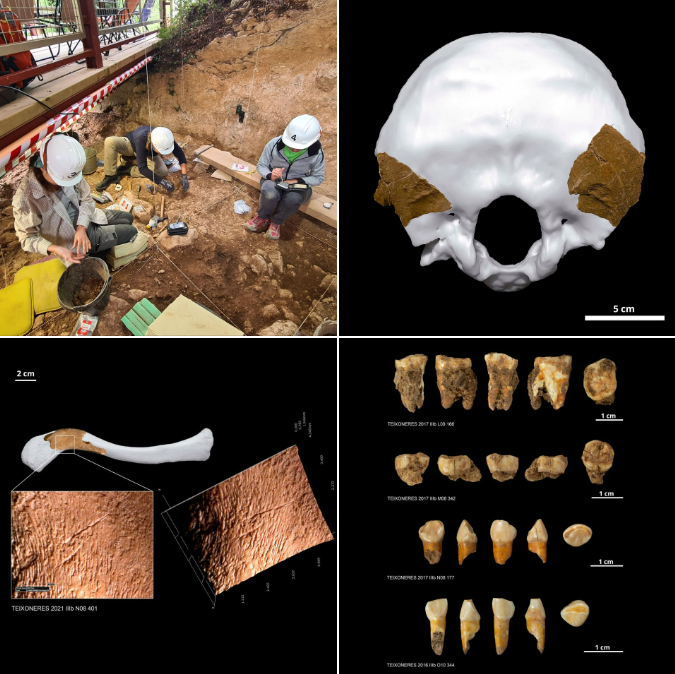
Restos neandertales de Cova de les Teixoneres y marcas de canibalismo. Crédito: IPHES (Institut Català de Paleoecologia Humana i Evolució Social)
- El estudio (preprint) del ADNmt de los restos neandertales de Mandrin (individuo apodado Thorin, datado en 50-42 ka) ha dado como sorprendente resultado que estaba genéticamente más próximo al neandertal de Forbes’ Quarry (Gibraltar-1, apodado Nada), que a otros neandertales más cercanos geográficamente. Esto indica un aparente aislamiento de ciertos grupos de una población fragmentada y tardía de neandertales, de la población mayoritaria de Europa occidental [+].

Restos del individuo neandertal apodado Thorin, de la cueva Mandrin (Francia). Crédito: L. Slimak et al (2023).
- Se ha estudiado el maxilar y la dentición del individuo infantil neandertal Guercy 3 (de unos 7 años al morir), datado en 120 ka, comparándolo con preneandertales (MIS 14–9), neandertales tempranos (MIS 7-5e), neandertales tardíos (MIS 5d-3), y con Homo sapiens del Paleolítico medio (MIS 5), Paleolítico superior (MIS 3–2), Paleolítico superior final (MIS 1) y recientes. La conclusión es que el maxilar de Guercy 3 se aleja de la morfología de los neandertales tardíos, en rasgos como el posicionamiento de la raíz del proceso cigomático, las regiones infraorbitarias y nasales, los alveolos premaxilar, bucal y labial, el seno maxilar, la cavidad nasal y la verticalidad de la dentición anterior. En cambio, se acerca a la morfología preneandertal de la Sima de los Huesos; y su dentición es típicamente neandertal [+].
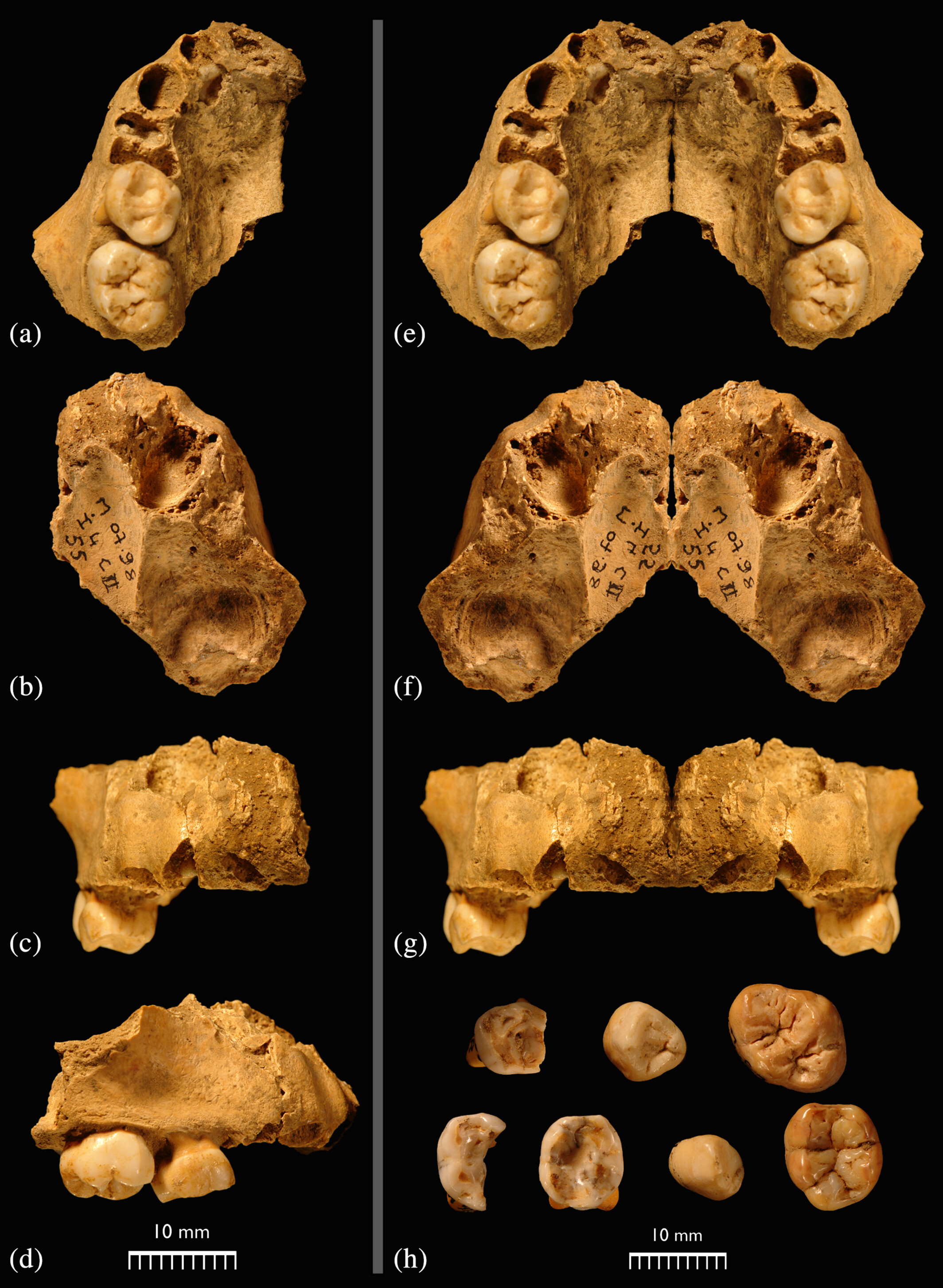
Individuo Guercy 3. Crédito: G. D. Richards et al (2023)
- Se ha analizado la composición de los conjuntos de fauna de mamíferos asociados a sitios neandertales y de humanos modernos en el rango 60-30 ka en la península ibérica. No se han hallado diferencias significativas entre ambas especies, aunque la regionalización bioclimática es más fuerte en los conjuntos asociados a humanos modernos, lo que puede indicar una diferencia en la duración de la ocupación del sitio o la movilidad de forrajeo entre ambas especies [+].
- Se han reconstruido los genomas bacterianos encerrados en la placa dental de 12 neandertales de 102–40 ka, 34 humanos modernos de 30 ka-150 a y otros 18 actuales, destacando el microbioma oral muy bien conservado de la llamada Dama Roja de El Mirón (Cantabria) de 19 ka. El objetivo fue recuperar información genética de bacterias antiguas, recrear sus funciones arcaicas introduciendo su ADN en bacterias modernas, y analizar la producción de compuestos similares a los que habrían hecho sus homólogas antiguas, para comprender mejor cómo interactuaban las bacterias entre sí o con sus huéspedes, y potencialmente encontrar nuevos compuestos biológicamente activos con actividades antibióticas [+].
- Los humanos actuales tenemos una herencia genética de los neandertales que da lugar a una mayor altura nasal, posiblemente como ventaja evolutiva que permitiría la adaptación de Homo sapiens al clima de Euroasia. A esta conclusión llega un estudio con más de 6000 individuos iberoamericanos, de ascendencia mixta europea, nativa americana y africana, de los cuales se analizó su información genética y las fotografías de sus rostros, observando las distancias entre puntos, como la punta de la nariz o el borde de los labios, e identificando la asociación de rasgos faciales con la presencia de diferentes marcadores genéticos. Se identificaron 33 regiones genómicas asociadas a la forma de la cara, 26 de las cuales se pudieron comparar con datos de individuos de otros orígenes de Asia oriental, Europa y África. En la región genómica ATF3 , muchos individuos con ascendencia nativa americana y de Asia oriental mostraban una selección positiva y de herencia neandertal relacionada con la mencionada altura nasal mayor [+].
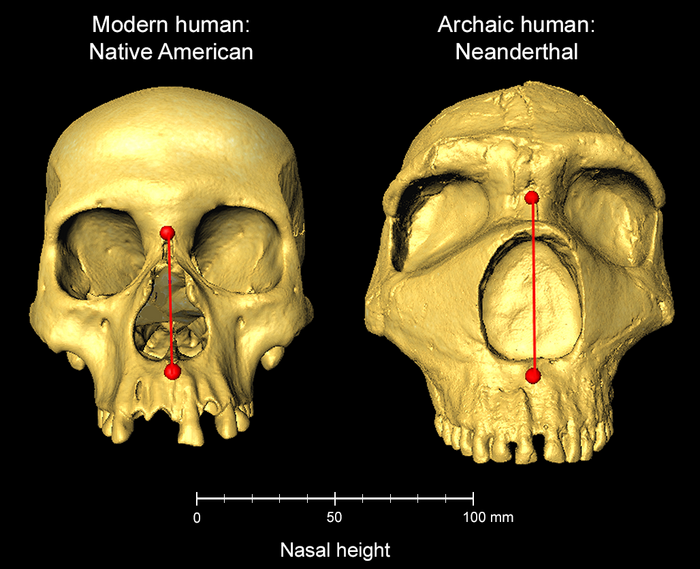
Altura nasal en humano moderno y en neandertal. Crédito: Kaustubh Adhikari, UCL
- A partir de las analogías de las industrias del Paleolítico superior inicial europeo con la secuencia contemporánea en sitios del Mediterráneo oriental, especialmente el de Ksar Akil (Líbano), se sugiere la existencia de tres oleadas distintas de humanos modernos por el interior de Europa, es decir, tres fases del primer Paleolítico superior levantino que son análogas a las tres industrias asociadas a dichas oleadas en Europa occidental: el Neroniano hace 50-60 ka, el Chatelperroniense hace 45 ka y el Protoauriñaciense hace 42 ka [+].
- Se ha presentado el objeto más antiguo para el trabajo del cuero (y probablemente de ropa) por cazadores-recolectores auriñacienses hace 39,6 ka, mucho antes de la introducción de agujas de hueso 15 ka después. Se trata de un hueso en Canyars (noreste de España) que se empleaba como soporte para perforar pieles de animales con buriles. Tiene 28 perforaciones con una distribución que sugiere el objetivo de dar una puntada lineal de, como mínimo, diez agujeros equidistantes, con una media de 5 mm entre cada uno. Asimismo, en la superficie ósea se observan otras cinco marcas de perforación, cada una de las cuales produce dos o tres agujeros [+].

Tablero con orificios para el trabajo del cuero. Crédito: Doyon et al (2023)
- Se ha documentado con diferentes métodos la utilización recurrente de la cueva de Nerja (sur de España) por humanos durante unos 35 ka. La datación del C14 de 53 carbones identificó 12 fases de visitas prehistóricas a la cueva entre 41 ka y 3 ka (cal BP), retrocediendo 10 ka el origen de la ocupación humana de este sitio. El periodo 8003–2998 cal BP contiene al menos 64 incursiones distintas, con un promedio de una visita cada 35 años para el Neolítico. Según el análisis espacial, no todas las áreas de la cueva fueron utilizadas en los mismos momentos, destacando la repetición de visitas a ciertos sectores específicos de las galerías inferiores de la cueva [+].
- Una nueva técnica permite extraer el ADN antiguo de humanos que han manipulado objetos de hueso. En concreto, se ha hallado ADN de una mujer en este colgante de diente de ciervo de unos 20 ka procedente de la cueva Denisova. Se suma a las técnicas para estudiar el ADN recuperado del suelo o de los sedimentos, que nunca dejan de asombrar [+].
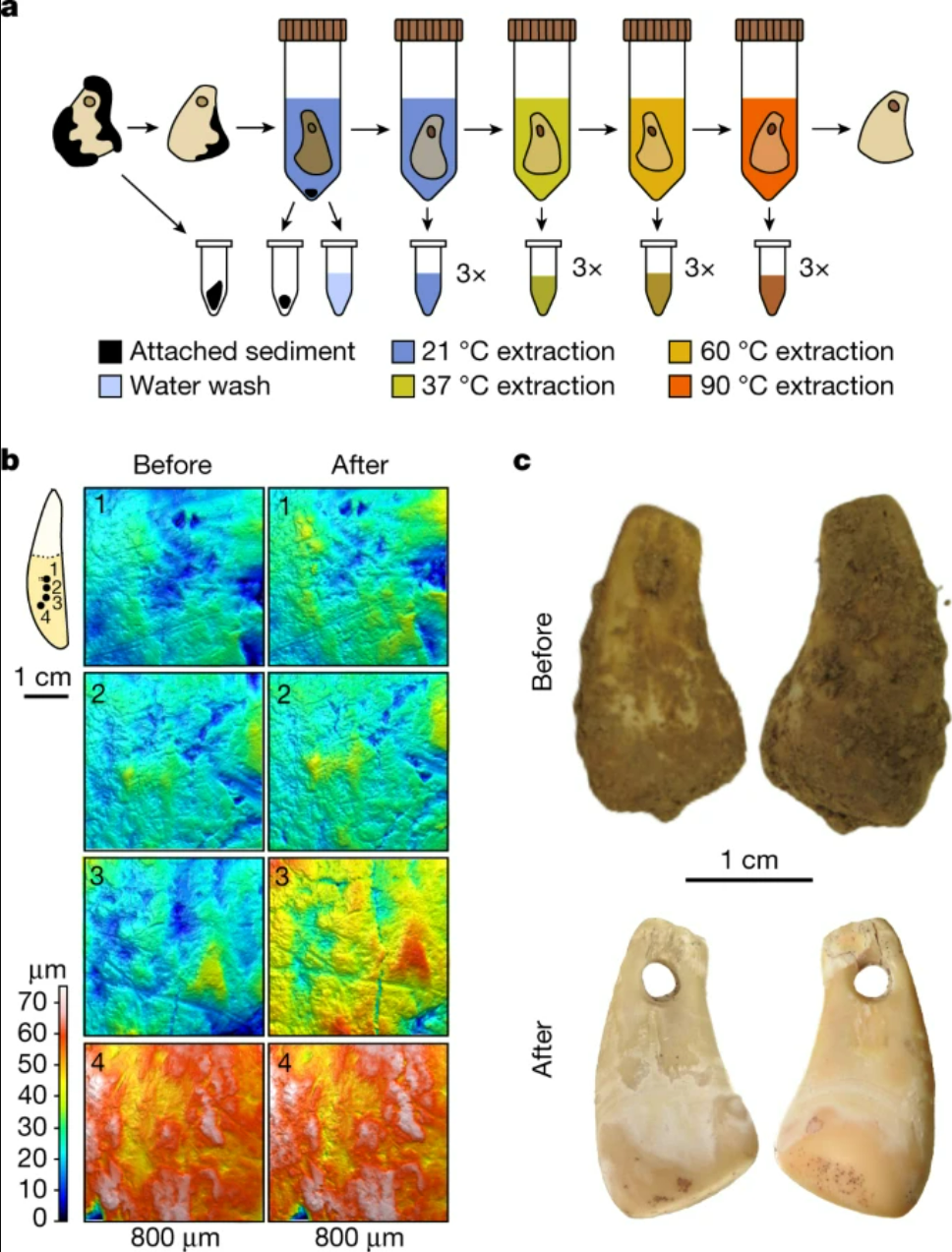
Colgante de cueva Denisova. Crédito: Essel, E., Zavala, E.I., Schulz-Kornas, E. et al. (2023).
- Una inusual acumulación y distribución de piedras sugiere el uso de este espacio de forma separada de la cueva El Mirón (norte de España) posiblemente como zona dedicada a la preparación de herramientas óseas en el Magdaleniense inferior [+]. Es excepcional, debido al escaso número de espacios de este tipo conocidos hasta la fecha.

The rock alignment in Level 115: top) during excavation (photograph by L.G. Straus. From left to right are squares U9, U8 and the north half of U7—that is, the eastern portion of the feature); bottom) in plan view (see Straus & González Morales Reference Straus, González Morales, Valde-Nowak, Sobczyk, Nowak and Źrałka2018: fig. 2).‘Éboulis’ indicates angular limestone spall (plan by L.G. Straus and R.L. Stauber).
- Entre una colección de 1100 huesos de ave recogidos en el yacimiento natufiense de Eynan-Mallaha (Israel) en los años 50, se ha descubierto que seis de ellos son flautas, una de ellas intacta, que mide 6,5 cm. Tienen unos 12.000 años. Al tocarlas, las flautas emitían un sonido agudo similar al de algunas aves rapaces, como se puede comprobar en el siguiente vídeo [+].
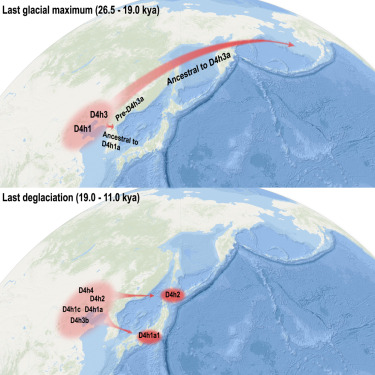
- Aunque existen distintas evidencias genéticas de que los antepasados de los primeros nativos americanos proceden principalmente de Siberia, existen otras fuentes ancestrales que también contribuyeron. Así, se han identificado dos eventos de radiación y dispersiones de ascendencia matrilineal desde la costa norte de China: uno hasta América durante el Último Máximo Glacial (26,5-19 ka) y otro hacia Japón hace 19-11 ka [+].
- Hace 9000 años, se realizaron en Jordania y Arabia Saudí un conjunto de grabados que representan con extrema precisión megaestructuras neolíticas vecinas, indicando una maestría mental de la percepción espacial inesperada para esa antigüedad [+].

(A) Desert kite plan from Kazakhstan (Ustyurt Plateau). (B) Desert kite plan from Armenia (Mount Aragats). (C) Desert kite plan from Jordan (Harrat al-Shaam). (D) Desert kite plan from Saudi Arabia (Khaybar). (E) Distribution area of desert kites from western Arabia to Uzbekistan. (F) Oblique aerial picture of a desert kite in Jordan (Harrat al-Shaam, photo OB, Globalkites Project). (G) Oblique aerial picture of a desert kite in Saudi Arabia (Khaybar, photo Khaybar Longue Durée Archaeological Project, RCU-AFALULA-CNRS). (H), (I), (J) Desert kite pit-traps during and after excavation (of half the pit) in the Harrat al-Shaam region of Jordan (photos RC, OB, WAA, Globalkites Project).
- En la transición neolítica al estilo de vida agrícola en el noroeste de África, se observa una notable continuidad genética desde el Paleolítico superior hasta el Neolítico inicial (entre 15 y 7,5 ka), en la que los grupos de cazadores-recolectores adoptaron prácticas agrícolas aprendidas desde un grupo de individuos de ascendencia neolítica mayoritariamente europea (que, a su vez, se remonta al Neolítico de Anatolia) con alrededor de un 20% de ascendencia local. Es decir, eran probables descendientes de agricultores inmigrados recientemente, y por tanto, la agricultura fue introducida por emigrantes europeos y adoptada rápidamente por los grupos locales. En el Neolítico medio (6,7-6,2 ka), coincidiendo con la llegada del pastoreo a la región, aparece en el Magreb otra ascendencia de origen levantino que probablemente siguió la costa norteafricana. Las tres ascendencias se mezclaron durante el Neolítico final. Estos cambios de ascendencia en la neolitización del noroeste de África probablemente reflejaban un paisaje económico y cultural heterogéneo, en un proceso más polifacético que el observado en otras regiones [+].
Resúmenes de trimestres anteriores
- Paleoantropología 2023 novedades por trimestre: 1T
- Paleoantropología 2022 novedades por trimestre: 1T | 2T | 3T | 4T
- Paleoantropología 2021 novedades por trimestre: 1T | 2T | 3T | 4T
- Paleoantropología 2020 novedades por trimestre: 1T | 2T | 3T | 4T
- Paleoantropología 2019 novedades por trimestre: 1T | 2T | 3T | 4T
- Paleoantropología 2018 novedades por trimestre: 1T | 2T | 3T | 4T
- Paleoantropología 2017 novedades por trimestre: 1T | 2T | 3T | 4T

Gracias Roberto, es delicioso leer estos resúmenes trimestrales que haces. Un cordial saludo
Me gustaLe gusta a 1 persona
Gracias por comentarlo, anima mucho a seguir.
Me gustaMe gusta
Felicitaciones, como siempre, extraordinaria información sobre evolución y Paleoantropología.
Me gustaLe gusta a 1 persona
Los resúmenes son una puerta a buscar más info y a emocionarse. Gracias.
Me gustaLe gusta a 1 persona
Cómo me alegro. Gracias por compartirlo.
Me gustaMe gusta