
Sin llegar a las 32 del trimestre anterior, pero este tercer trimestre del año no se queda atrás en cuanto a novedades publicadas en esta apasionante ciencia. Aquí va el resumen de 27 de ellas:
- Comenzamos por MRD VP-1/1, el primer cráneo casi completo conocido de Australopithecus anamensis, y primera vez que podemos poner cara a un miembro de esta especie. El análisis de su morfología parece confirmar la posición filogenética de Au. anamensis entre Ardipithecus ramidus y Au. afarensis, aunque la datación de los materiales de Au. anamensis y Au. afarensis pone de manifiesto que ambas especies tal vez coexistieron unos 100.000 años, y pone en duda que la primera dio lugar a la segunda por anagénesis [+info].

Cráneo MRD y reconstrucción. Crédito: The Cleveland Museum of Natural History
- Recordemos a Little Foot (Stw 573), el esqueleto más completo de Australopithecus, hallado en Sterkfontein tras un laborioso trabajo de extracción de la roca cementada durante muchos años. El estudio de los huesos largos de Little Foot concluye que las proporciones brazos/piernas coinciden con las de Lucy, y por tanto descarta una explicación alométrica para el elevado índice humerofemoral de Lucy, como en ocasiones se ha planteado. En vez de eso, el estudio propone una evolución de las extremidades en dos fases: 1) un modesto aumento de la longitud de las piernas y un acortamiento simultáneo del antebrazo en Ardipithecus y Australopithecus ; 2) un considerable alargamiento de las piernas junto con una disminución de los brazos en Australopithecus y Homo sapiens [+info].
- El análisis de las acumulaciones cíclicas de litio en dos especímenes infantiles de Australopithecus africanus de Sterkfontein, indica que se alimentaban casi exclusivamente de la leche materna hasta los 6-9 meses después de nacer, y después del año nuevamente en las estaciones secas ante la ausencia de alimentos. El periodo de lactancia en esta especie es más comparable al de los humanos modernos, que al de los chimpancés actuales (más largo) [+info].
- Un conjunto de herramientas de la formación Dawqara en Zarqa Valley (Jordania), datadas en 2,48 Ma, constituyen la presencia más antigua de homininos fuera de África [+info]. Este hallazgo se une otras evidencias humanas sorprendentemente antiguas que hemos conocido en meses anteriores: las herramientas de Ain Boucherit (Argelia) de 2,4 Ma y las de Shangchen (China) de 2,12 Ma.
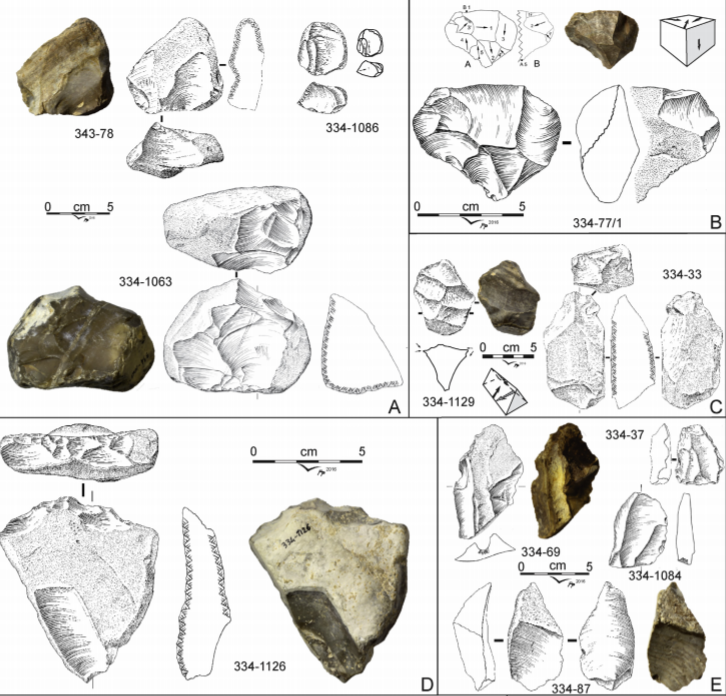
Dawqara. Credit: Scardia G et al (2019)
- En los años 80, en Okote Member (Koobi Fora, Kenia) se encontraron pequeñas áreas de tierra enrojecida asociadas a herramientas de 1,5 Ma. Mediante técnicas de susceptibilidad magnética y luminiscencia se demostró que el coloramiento fue provocado por el fuego, pero se discute sobre si fue natural o provocado por la acción humana. El reestudio de los materiales de Okote en 2019 confirman la datación, constituyendo las evidencias más antiguas de uso del fuego por humanos [+info].
- Mediante un nuevo algoritmo bayesiano se ha estimado la probable cantidad de introgresiones de antepasados de los humanos modernos existentes a lo largo del genoma de los neandertales y denisovanos. Pues bien, esto ocurre en el 3% del genoma neandertal (y el flujo genético sucedió hace 200-300 ka), y en el 1% de las regiones del genoma denisovano (y el 15% de las regiones se han pasado a los humanos modernos a través del flujo genético) [+info].
- La hemimandíbula denisovana de Xiahe, de 160 ka, publicada en mayo 2019, tiene un molar con 3 raíces, un rasgo que solo ocurre en el 3,5% de los humanos no asiáticos pero en más del 40% en poblaciones de China y del Mar de Bering. Se sugiere que este rasgo puede ser arcaico y surgir en los humanos modernos como consecuencia de la introgresión de los denisovanos [+info].

The 3-rooted lower molar anomaly. Source: Bailey, Hublin & Antón (2019)
- Los antepasados de los humanos modernos se hibridaron con al menos 5 grupos arcaicos distintos, 2 de las cuales son conocidos (neandertales en el oeste euroasiático y denisovanos en Altai). La localicación de los otros eventos de hibridación se situarían en el sureste asiático/norte de India y en el este: Filipinas, Sunda y Flores [+info].
- Fueron noticia dos preciosos grabados hechos sobre hueso hace 120-125 ka encontrados en Lingjing (provincia de Henan, China), uno de ellos con restos de ocre sobre las incisiones [+info].

Two engraved bone fragments from Lingjing, China. Credit: F. d’Errico & L. Doyon
- Un estudio ha observado que la mitad de 23 neandertales analizados padecían exóstosis en el conducto auditivo externo, de forma similar a los humanos que practican deportes acuáticos o viven en ambientes húmedos fríos («oído del surfista»). ¿Pasaban tiempo en el agua para pescar y recolectar marisco y moluscos? (lo cual no se observa en el registro arqueológico ni en los isótopos de los dientes) [+info].
- Adicionalmente, la reconstrucción de la estructura de la porción cartilaginosa de la trompa de Eustaquio de los neandertales indica que estas regiones en los neandertales eran estrechas y horizontales, parecidas a las de los infantes actuales. Esto sugiere que padecerían infecciones del oído medio de manera frecuente [+info].
- Entre 10 y 14 individuos neandertales, la mayoría niños, dejaron 257 huellas sobre la arena cerca de la costa de Le Rozel en Normandía (Francia) hace 80 ka [+info].

Una de las 257 huellas. Crédito: Dominique Cliquet
- Se han publicado nuevas evidencias del empleo de adhesivos por los neandertales, hallados como restos en varias herramientas de distintos tipos en las cuevas de Fossellone y Sant’Agostino (Italia), con datación 55-40 ka. Dichos restos consistían en resina de coníferas calentada brevemente para moldearla, y en una de las herramientas se encontró también cera de abeja mezclada con la resina [+info].
- Se ha conseguido analizar el ADN nuclear de dos especímenes neandertales europeos de 120 ka, de los cuales solo se había conseguido ADN mitocondrial: un fémur de la cueva Hohlenstein-Stadel (Alemania) y un maxilar de la cueva Scladina (Bélgica). Pues bien, su genoma se parece más al de los últimos neandertales europeos de hace 40 ka (Vindija, Croacia) que a los neandertales de Denisova de cronología similar a la suya, indicando una continuidad genética de esta especie durante 80 ka [+info].

Sites from which partial to complete nuclear genomes from Neandertals (or their ancestors in Sima de los Huesos) were retrieved. Credit: Stéphane Peyrégne et al. (2019) Sci Adv 2019;5:eaaw5873
- Una nueva técnica aísla la contaminación moderna en el ADN antiguo previamente a su secuenciación, y se ha aplicado a los cráneos neandertales de Gibraltar. El resultado es la confirmación de que el cráneo de Forbes’ Quarry era de una mujer infantil y el infantil de Devil’s Tower era de un niño, y que el adulto era genéticamente más similar a neandertales más antiguos euroasiáticos que a especímenes peninsulares más recientes [+info].
- Se ha encontrado la primera evidencia de presencia humana en alta montaña en África, en concreto en las montañas de Bale en el sur de Etiopía (refugio de Fincha Habera, a 3500 m de altitud) hace entre 31 y 47 ka. Habitaron aquellas altas llanuras durante épocas largas aprovechando los periodos secos, cuando existirían abundantes recursos hídricos y fauna pequeña (roedores). Hace 10 ka aquel lugar fue habitado de nuevo. Hoy los habitantes de esa región muestran una adaptación a la vida en altura [+info].
- El cráneo Apidima 1 (Grecia), con datación 210 ka, supondría la presencia más antigua de Homo sapiens en Eurasia. La pertenencia a nuestra especie parece apoyada por su morfología: es corto, tiene un contorno mediosagital redondeado y carece del típico moño occipital neandertal. También posee algunas plesiomorfias que comparte con otros especímenes del Pleistoceno Medio, como la forma de las paredes parietales [+info].

The fossil crania of Apidima 2 (up) and Apidima 1 (bottom). Credit: Harvati K et al (2019)
- La desaparición del oso de las cavernas (Ursus spelaeus) no fue lenta, sino que su población se mantuvo estable durante milenios, desde que hace unos 450 ka un ancestro común dio lugar a cinco linajes principales de osos cavernarios que se dispersaron por Europa, comenzando a decaer hace 50 ka y acelerándose bruscamente su extinción hace 40-35 ka, según un estudio de su ADN mitocondrial [+info].
- La revisión de una la mandíbula humana infantil de la cueva El Castillo (España), junto con la reinterpretación de las notas de Obermaier y Vallois que la estudiaron inicialmente, ha dado una datación de unos 27 ka y la presencia de marcas de corte para extraer la lengua, indicando un posible acto funerario o de canibalismo, comportamientos que no son frecuentes en otros esqueletos del Gravetiense [+info].

Mandíbula infantil de El Castillo. Crédito: Pedro Saura/Raquel Asiaín
- Nuevos hallazgos enriquecen el conocimiento sobre los primeros poblamientos de América. En este caso, el conjunto de herramientas y restos de fauna procesada hace 16,5 ka en el sitio de Cooper’s Ferry, a orillas del río Idaho, reforzando la hipótesis de una llegada por la costa del Pacífico más de un milenio antes de que se abriera un pasillo de hielo por el interior [+info].
- También en cuanto a la colonización de Sahul (plataforma continental que unió Australia y Nueva Guinea), dos nuevos estudios plantean que al menos 130 humanos fueron capaces de navegar de forma planificada por una ruta norteña hace unos 50 ka [+info].
- Mediante un modelo estadístico se ha estimado cómo pudo haber sido la morfología del cráneo virtual del «primer humano moderno». Este cráneo virtual presenta una mayoría de rasgos que se considerarían modernos (forma globular, frente alta, grandes parietales, base del cráneo curvada, cara retraída, con cigomáticos angulados y proyectados, fosa canina bien marcada..) pero también algunos rasgos más arcaicos (frontal ligeramente retrocedido, arcos supraorbitales proyectados, prognatismo alveolar de la cara, ligero moño occipital…) [+info].
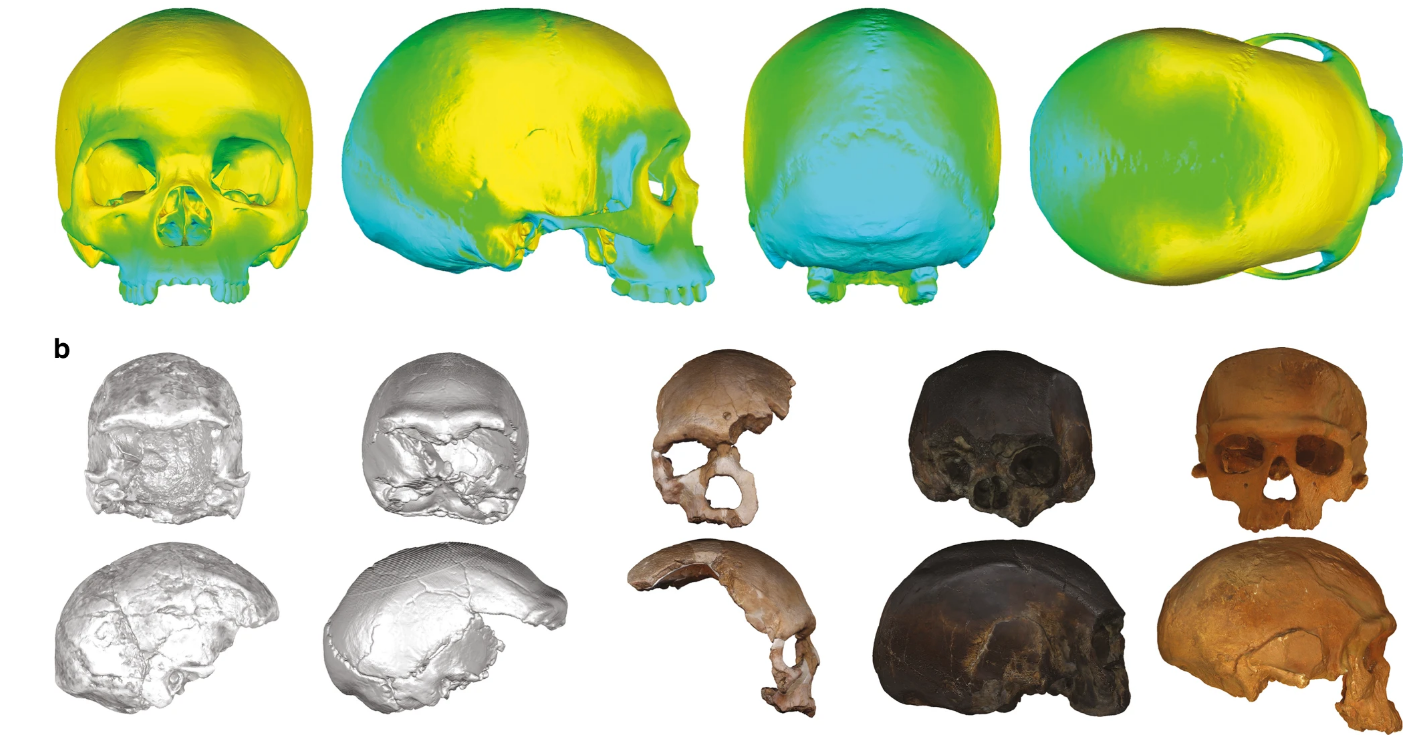
Arriba: Cráneo virtual vLCAs (azul=rasgo mayor en vLCA1, amarillo=mayor en vLCA2). Abajo: cráneos Omo II, LH18, Florisbad, KNM-ES 11693, Irhoud 1. Fuente: Mounier & Mirazón Lahr (2019)
- Hace un año, un estudio en Cell que abarcaba la morfología de los primeros humanos modernos, la evidencia genética, el despliegue cultural de los distintos grupos en África, y el entorno ecológico en que se desarrollaron, planteaba una evolución en mosaico de distintas poblaciones humanas que interactuaron de forma variable en el tiempo, lo que dio lugar a nuestra especie Homo sapiens [+info]. Ahora los autores han publicado el estudio revisado en Nature, en que los sucesivos estudios genéticos parecen reforzar un modelo metapoblacional para el origen panafricano de nuestra especie, a partir del flujo genético entre poblaciones primitivas y dispersas por todo el continente [+info].
- En el yacimiento israelí de Qesem Cave, aves como el cisne, paloma, cuervo y estornino no solo fueron aprovechadas como alimento durante el Pleistoceno Medio, sino también por sus plumas con una finalidad no nutricional (tal vez simbólica), según muestran las marcas de corte encontradas [+info].

Crédito: Ruth Blasco et al (2019)
- A partir de la información genética contenida en el fósil Denisova-3, la falange de un dedo meñique, se ha aplicado una nueva técnica que asocia cambios en la actividad de regulación genética en fósiles (metilación del ADN) con cambios anatómicos entre grupos humanos, con el objeto de predecir la apariencia física de los denisovanos. Asíi, se han determinado 56 rasgos que los distinguen de los neandertales y de los humanos modernos, 34 de ellos en el cráneo [+info].
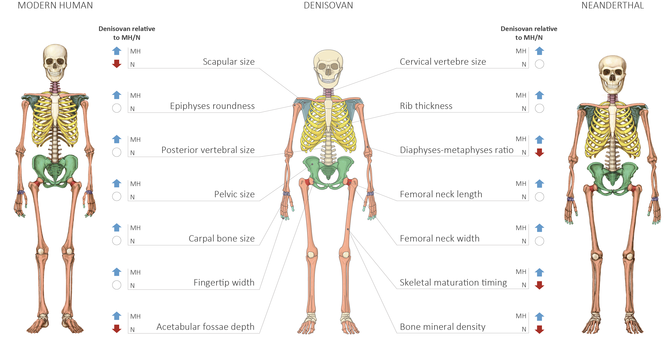
Modelo anatómico de un humano moderno, un neandertal y un denisovano. Crédito: Maayan Harel
- El estudio de la pelvis de Rudapithecus hungaricus, un homínido que vivió hace 10 millones de años en la actual Hungría, sugiere que la capacidad de bipedación pudo surgir de alguna forma de locomoción más antigua de lo que suponemos. Aunque aquel ser tenía vida arbórea, su espalda baja era flexible, y en el suelo podría haber tenido la capacidad de estar erguido como los humanos. La hipótesis planteada es que, que si los humanos evolucionaron de una estructura corporal similar a la de un simio africano, se habrían requerido cambios sustanciales para alargar la espalda baja y acortar la pelvis; pero si evolucionaron de un antepasado más parecido a Rudapithecus, esta transición habría sido mucho más directa [+info].
- La evidencia más antigua de ocupación humana en Francia es de 650-670 ka, que es 150 ka más antigua que la que se conocía anteriormente, gracias al hallazgo de 260 artefactos líticos (entre ellos 5 bifaces) en el yacimiento de Moulin Quignon al norte [+info].
Más información:
