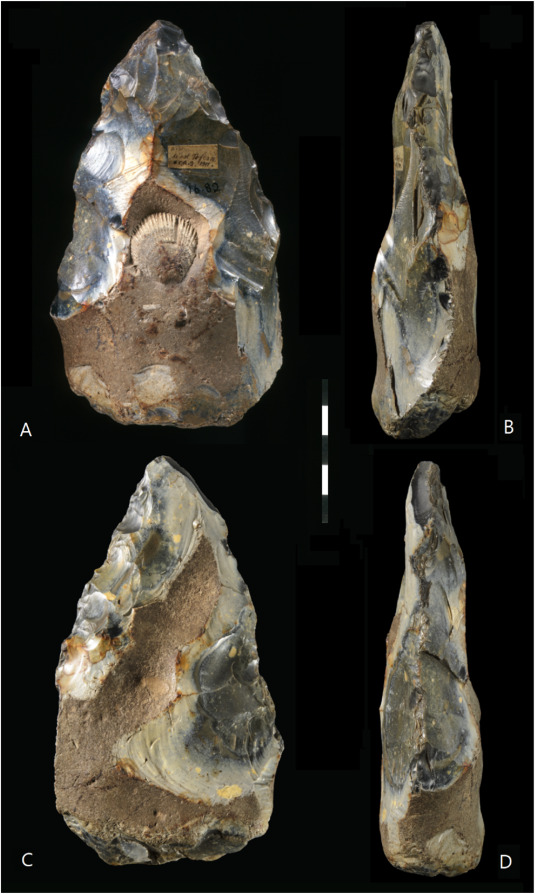
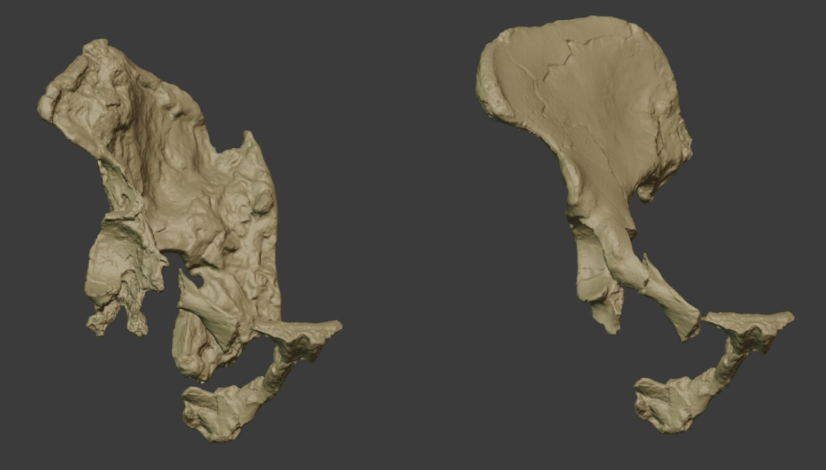
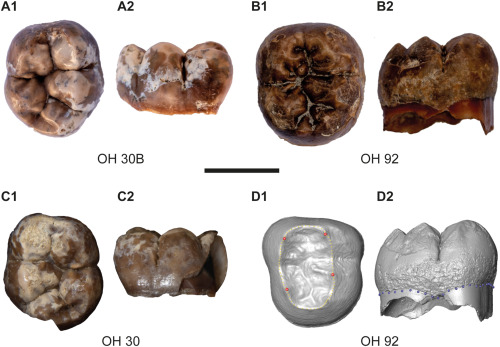
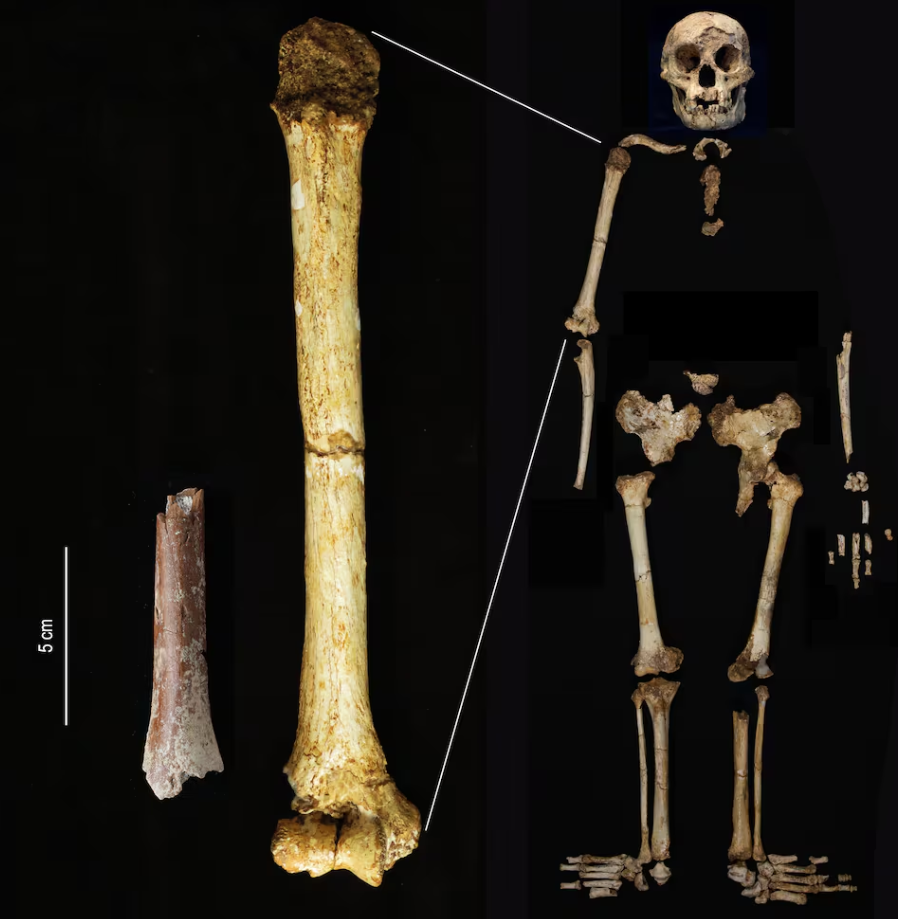
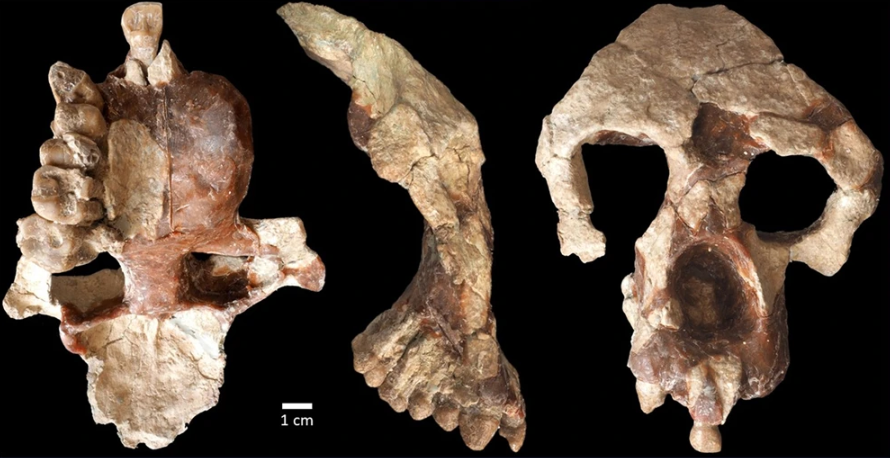
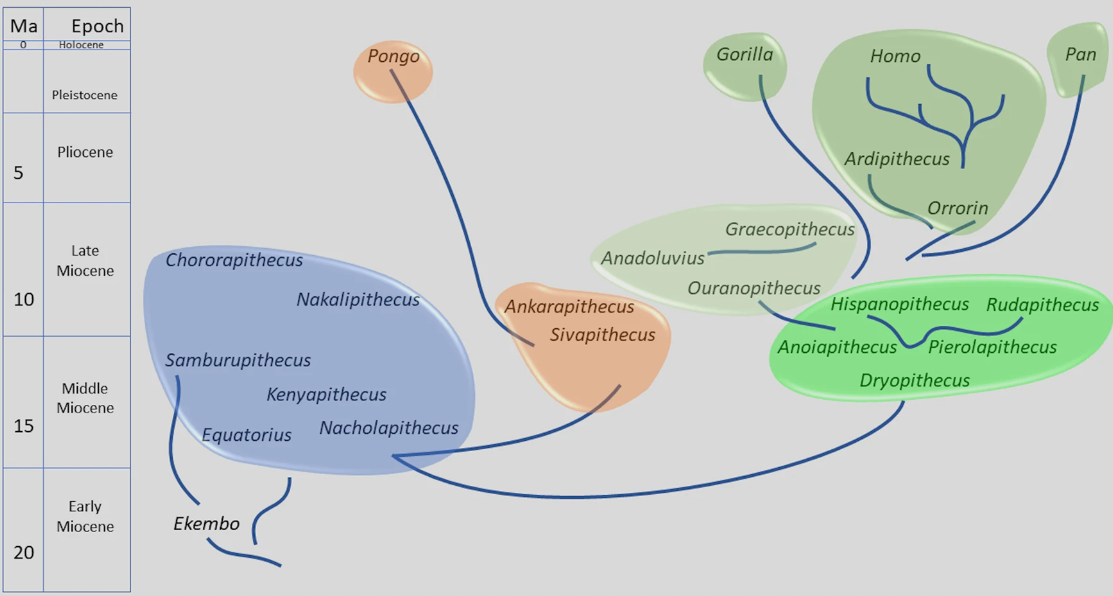
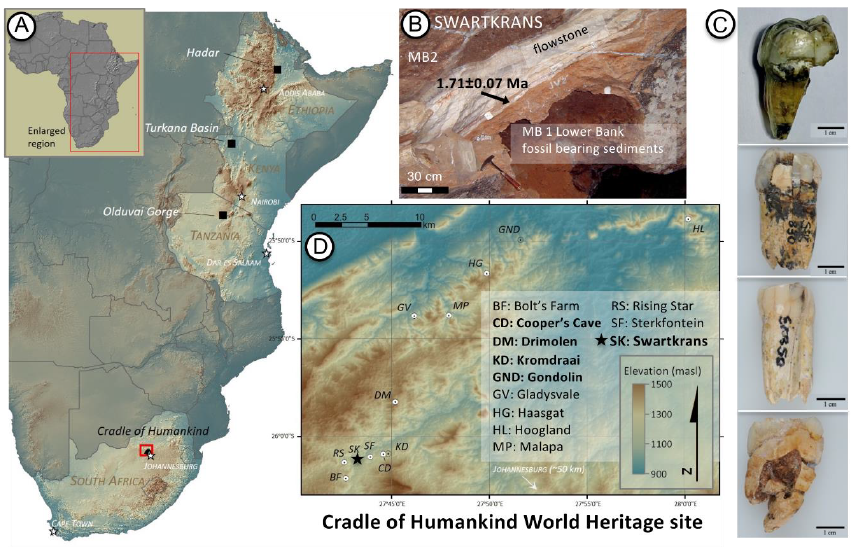
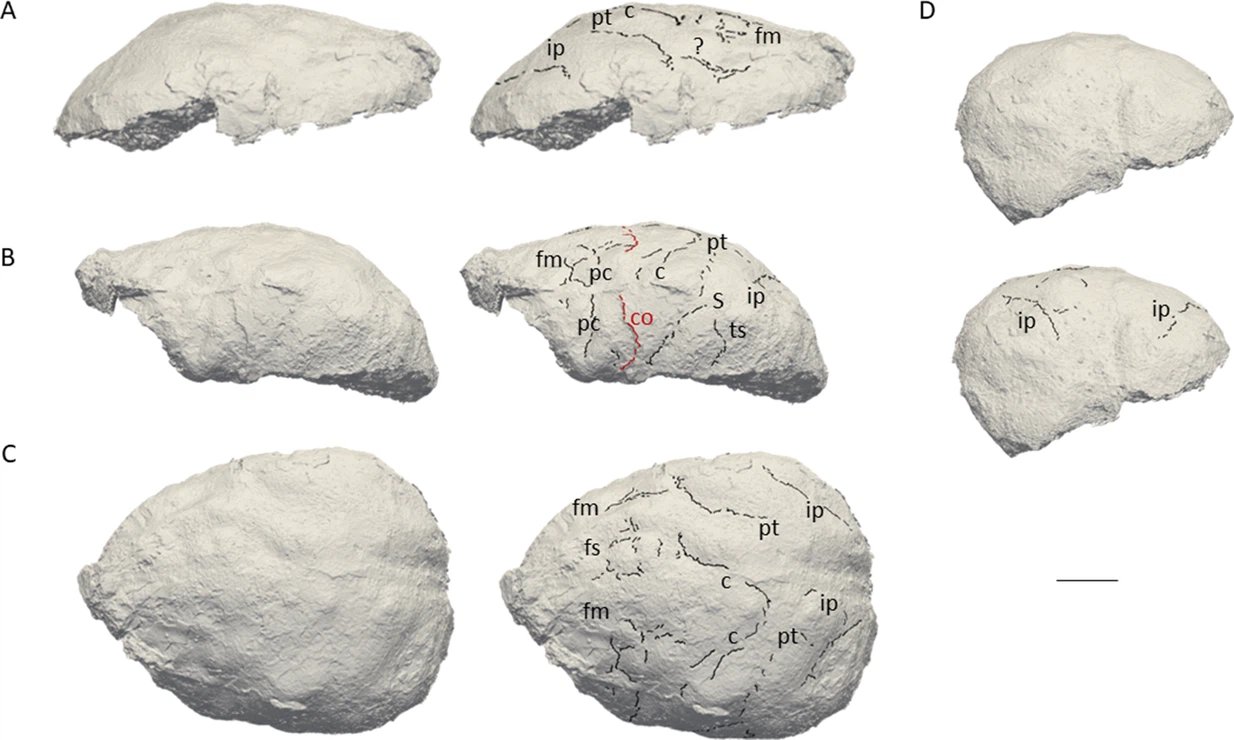
Cerramos un gran año en hallazgos paleoantropológicos. En el último trimestre recopilé 46 publicaciones que ilustran la riqueza acumulada: los cuatro resúmenes trimestrales de 2025 suman 198 novedades. ¡Feliz 2026!
- Combinando modelos tridimensionales de cráneos y métodos comparativos filogenéticos, Gómez-Robles y colaboradores han analizado la velocidad a la que distintas regiones del rostro y del neurocráneo cambiaron a lo largo del tiempo en distintos linajes de primates. El linaje humano duplicó la tasa esperada de cambio morfológico, y esto sucedió en la mayoría de los rasgos craneofaciales, separándose claramente de otros grandes simios y mostrando una selección direccional.
- Al contrario que el desarrollo prolongado en el neurocráneo de Homo sapiens y Homo erectus, en Australopithecus afarensis y Paranthropus robustus los patrones de desarrollo son variables (Braga, Alemseged y Gilissen, 2025). De hecho, Au. afarensis exhibe un crecimiento endocraneal prolongado en comparación con P. robustus, un contraste incluso mayor que el de Pan paniscus y Pan troglodytes, lo que sugiere que el desarrollo endocraneal en Au. afarensis se asemeja más al de H. sapiens que al de P. robustus.
- El análisis 3D morfométrico de las inserciones ligamentarias radiocarpianas (Casado et al., 2025) en 110 radios de homininos fósiles aporta información sobre sus patrones locomotores: Au. afarensis, Au. anamensis y Au. sediba combinaban locomoción bípeda y arbórea similar a chimpancés y orangutanes; P. robustus tenía una locomoción terrestre más similar a la de los gorilas; neandertales y H. sapiens basales muestran una anatomía similar a la de los humanos actuales, pero algo más de robustez orientada a cargas manuales (caza/herramientas).
- El astrágalo ARA-VP-6/500-023 de Ardipithecus ramidus presenta similitudes con los astrágalos de chimpancés y gorilas, adaptados a la escalada vertical y al cuadrupedismo plantígrado terrestre. Además, el fósil tiene rasgos derivados consistentes con un mecanismo de impulso mejorado en el pie de Ar. ramidus. Prang y colaboradores proponen que el último ancestro común de humanos y chimpancés no sería un simio arborícola generalizado, sino un ancestro africano similar a un simio con un repertorio postural que incluía el cuadrupedismo plantígrado terrestre y la escalada vertical, del cual Ar. ramidus representaría un estadio intermedio, con modificaciones incipientes hacia una mayor eficiencia en bipedación. Por otro lado, la bipedación de Ardi sería distinta a la que emplean los grandes simios: su lateral del pie funcionaba mejor para el impulso lateral, dándole un empuje más eficiente al caminar.