<English version below>
Dos estudios publicados recientemente contribuyen de manera notable al (confuso) conocimiento que tenemos sobre la evolución humana durante el Pleistoceno Medio Europeo.
En 2010, Aida Gómez-Robles estudió la colección de dientes de los preneandertales de la Sima de los Huesos (SH) en Atapuerca. Tan solo 9 años después, se conoce mucha más información sobre aquellos humanos, principalmente: 1) La datación de los fósiles, establecida en unos 430 ka (Arnold LJ et al, 2014). 2) Gracias a la paleogenética, la confirmación de la relación de los homininos de SH con el linaje neandertal, lo cual ya indicaban muchos rasgos anatómicos. 3) Los dientes de SH son muy derivados, tienen un aspecto muy alejado del que se esperaría para un neandertal temprano, o una especie próxima al ancestro común de neandertales y humanos modernos. De hecho, algunos rasgos como la reducción del tamaño y la simplificación de la estructura se corresponden con neandertales muy posteriores, en el entorno de hace 100 ka.
Gómez-Robles utiliza una serie de modelos cuantitativos para calcular el rango temporal en que los homininos de SH pudieron desarrollar esa morfología dental, asumiendo que la tasa a la que esos neandertales tempranos evolucionaron sería constante y similar a la observada en otros homininos. La autora considera que, aunque es posible que los de SH evolucionaran mas rápido, eso no es lo observado en muchas otras especies de homininos. Incluso en especies como los parántropos, con una dentición muy derivada, esa tasa evolutiva es muy homogénea. Considerando dicha tasa, la divergencia entre neandertales y humanos modernos ocurriría hace un mínimo de 800 ka.
Sin embargo, el estudio del genoma neandertal indica una divergencia del linaje sapiens alrededor de hace 600 ka. ¿Qué implicaciones tiene la diferencia entre ambas estimaciones? Por una parte, los estudios genéticos tienen también una serie de asunciones como la tasa de mutación (indicativa de la rapidez y consistencia de los cambios genéticos acumulados a lo largo del tiempo), que pueden hacer variar los resultados de manera significativa. Por otra parte, el estudio anatómico puede estar indicando que la diferenciación morfológica (la divergencia en cuanto a fenotipos dentales de neandertales y sapiens) precede a la separación genética de ambas especies, según indica Gómez-Robles. Es decir, su anatomía indica que esos grupos ya estaban en direcciones evolutivas distintas, hay patrones de divergencia muy claros antes de la separación genética que, además, sabemos que tampoco fue total (hubo hibridaciones).
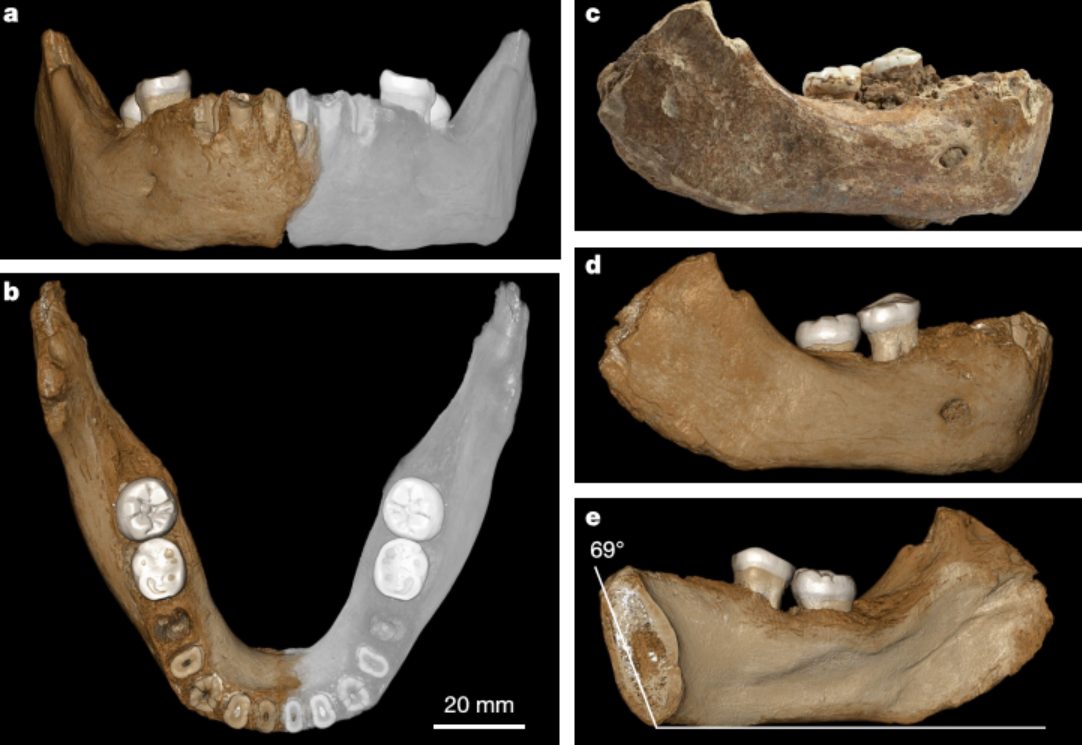
En el segundo estudio a que me refiero, Antonio Rosas y otros colegas plantean que la morfología craneal de los homininos del Pleistoceno Medio Europeo responde a una misma especie. Por tanto, hay una unicidad dentro de la diversidad que existe en las caras y en las mandíbulas de los homininos de Arago, Mauer, SH, Montmaurin… Para los autores, buena parte de esa diversidad puede explicarse por factores que vemos en nuestra especie y en otros simios, que se traducen en individuos con las caras más largas (dolicofacial) y otros con las caras más anchas (braquifacial).
Por tanto, a la vez que había grupos presapiens evolucionando en África, imaginamos que también habría preneandertales evolucionando en Europa, pero nos seguimos preguntando por los detalles detrás de esa dinámica evolutiva… Cuántos grupos humanos habitaron Europa durante el Pleistoceno Medio, cuál era la relación entre ellos, si se mezclaron, cuáles se extinguieron y cuáles sobrevivieron, cuántas especies representan, y cuáles son sus especies antepasadas. ¿Representan una sola especie politípica, Homo heidelbergensis? ¿Su anatomía era efectivamente previa a la separación genética de Homo neanderthalensis y Homo sapiens? ¿Es Homo heidelbergensis una especie «nicho» exclusivamente europea, y Homo antecessor el antepasado de ambos linajes hace 1 millón de años? Seguiremos atentamente la evolución del conocimiento en este apasionante periodo.

A model resulting from the successive pairwise similarity inclusion analyses in which only one polytypic species of Homo is recognized in the Middle Pleistocene of Europe. Source: Rosas A et al (2019). Tempo and mode in the Neandertal evolutionary lineage: A structuralist approach tomandible variation. Quaternary Science Reviews, fig. 8.