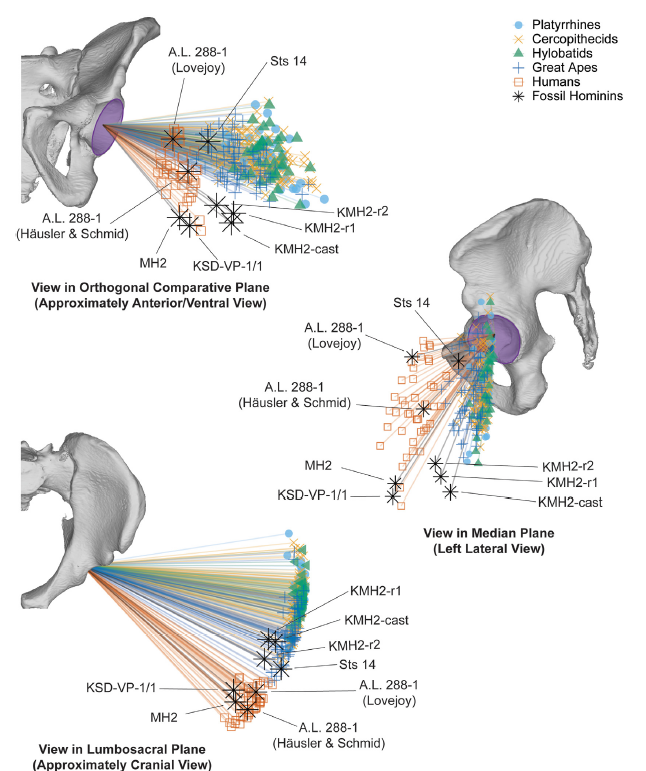
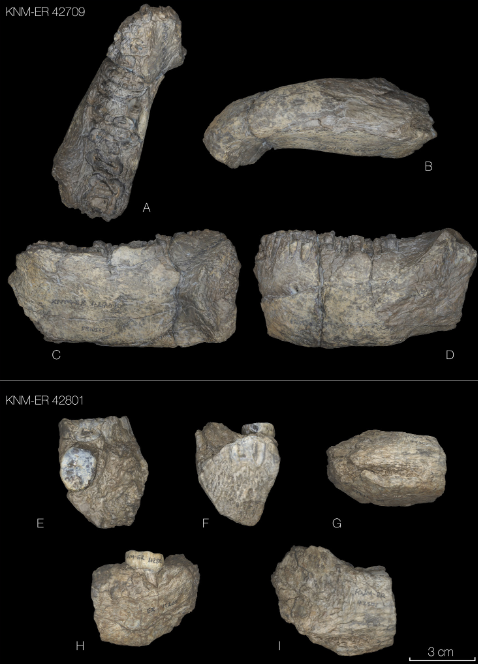
La comparación de las manos casi completas que se han podido recuperar de Australopithecus sediba (1,95 millones de años) y de Homo naledi (241-335 miles de años) permite observar cómo estas dos especies se adaptaron a sus entornos en el sur de África de distintos modos, equilibrando el uso de herramientas, el procesamiento de alimentos y la locomoción (Syeda et al., 2025).
Aunque ninguno de los dos homininos se ha asociado todavía directamente con herramientas de piedra, varios aspectos de la morfología de las manos y de las muñecas de ambos sugieren que poseían una destreza manual mucho más similar a la de los humanos que a la de los chimpancés o gorilas actuales. Es decir, aquellas manos probablemente manejaron herramientas.
En el caso de Homo naledi, cerca de la mano de uno de los cuerpos hallados en la antecámara Hill (contigua a la cámara de Dinaledi, en el sistema de cuevas Rising Star) se halló un artefacto de piedra que, según plantean en su preprint Berger y otros (2025), podría haber sido utilizado para realizar grabados parietales en forma de líneas cruzadas y formas geométricas. Aunque este objeto puede ser natural y no un producto de la talla de herramientas (Martinón et al., 2024), tampoco se descarta que H. naledi lo adoptara como útil. En todo caso, sorprende su asociación con el esqueleto, en contacto directo con el mismo.
Tanto H. naledi como Au. sediba son especies que conservan ciertos rasgos morfológicos de apariencia simiesca, sobre todo en las extremidades superiores. Estos rasgos les resultarían ventajosos para escalar o trepar, aunque no se puede determinar si realmente lo hacían, o tal vez esa morfología está reflejando los remanentes evolutivos de un ancestro escalador o trepador.
Sigue leyendo