
[English version below]
La paleoproteómica es una disciplina de secuenciación de proteínas procedentes de especímenes antiguos, que está comenzando a explotarse de manera más extendida en los últimos años. Permite trazar relaciones evolutivas comparando entre individuos las proteínas que cumplen la misma función, de forma similar a como hace la genómica comparando regiones genéticas equivalentes. Como las proteínas son más estables que el ADN y no se degradan tan rápidamente, en algunos materiales fosilizados muy antiguos sí se pueden recuperar y estudiar proteínas, pero no ADN. Por ejemplo, los restos humanos más antiguos de los que se ha podido analizar el genoma son los de Sima de los Huesos (Atapuerca), de 430 ka (miles de años), mientras que ya se han estudiado proteínas de otros restos humanos de casi 1 Ma (millón de años), y ya hay intentos con restos de 1,77 Ma. Por tanto, esta técnica viene a ayudar a donde la paleogenómica no llega. Veamos varios casos destacados:
- El cráneo de castor gigante Castoroides ohioensis. Datado en unos 12.000 años, fue encontrado en 1845 y estaba expuesto en el New York State Museum. El análisis publicado en 2016 de proteínas conservadas en los cornetes nasales, permite conocer la ubicación de esta especie dentro de Castoroidinae y su parentesco con otros representantes de esta subfamilia.
- También se ha publicado en 2019 el estudio taxonómico de distintas aves a partir de las proteínas de los huesos de especies extinguidas (dodo, Raphus cucullatus, y alca gigante, Pinguinus impennis), así como de otros especímenes más recientes de distintas especies.
- Un rinoceronte del género Stephanorhinus que vivía en Dmanisi hace 1,77 millones de años, conservaba proteínas en su esmalte dental que han ayudado a situarlo filogenéticamente como un grupo hermano del clado formado por el rinoceronte lanudo (Coelodonta antiquitatis) y el rinoceronte de Merck (Stephanorhinus kirchbergensis).
- Se ha podido confirmar el vínculo filogenético de Gigantopithecus con Pongo, gracias al estudio de seis proteínas preservadas en el esmalte y la dentina de un molar de Gigantopithecus blacki de 1,9 Ma encontrado en la cueva china de Chuifeng. Los linajes de Gigantopithecus y del orangután (su pariente actual más cercano) se separaron hace unos 10 Ma. Ese molar ha proporcionado los restos moleculares más antiguos que se han podido secuenciar de un fósil.
- También en 2019 se publicó una mandíbula denisovana de 160.000 años, hallada en en la cueva Karst Baishiya en Xiahe (China) a 3280 metros de altitud. Su pertenencia a dicha población humana se pudo determinar a partir de las proteínas de uno de sus molares.
- Finalmente, en abril 2020 se ha publicado el estudio proteómico de un diente de Homo antecessor, un primer o segundo molar inferior permanente (espécimen ATD6-92) de unos 800 ka, el proteoma más antiguo de un hominino que se ha podido presentar hasta la fecha, que viene a reforzar la ubicación de este taxón muy relacionado (grupo hermano) con el último ancestro común de sapiens, neandertales y denisovanos.
En este mismo estudio sobre Homo antecessor también se analizó un primer molar superior (D4163) de un hominino de Dmanisi datado en 1,77 Ma, sin resultados concluyentes, pero su antigüedad genera una enorme expectativa sobre la posibilidad de investigar las proteínas conservadas en el esmalte dental, el tejido más duro en el esqueleto de los mamíferos, y obtener grandes resultados para enriquecer nuestro conocimiento sobre las relaciones filogenéticas en la evolución humana.
Abusando del tópico, esto es solo el comienzo… Los resultados satisfactorios que proporciona la paleoproteómica para la revisión de nuestra filogenia, está ya empujando a investigadores a explotar más esta disciplina. Uno de los retos será averiguar por fin qué fue Homo erectus, taxón que abarca una diversidad de especímenes procedentes de grandes regiones de Asia y África (si incluimos a Homo ergaster) y una enorme línea temporal de más un millón y medio de años.
He consultado a José María Bermúdez de Castro su opinión sobre si será aplicable la paleoproteómica sobre algún otro resto de Homo erectus, y si podríamos esperar una relación entre los erectinos asiáticos y Homo antecessor:
Estoy convencido de que ya se están haciendo análisis con otros dientes, y H. antecessor entrará en la comparativa. Los próximos años serán divertidos y habrá más de una sorpresa. Ya se encontraron proteínas en un rinoceronte de Dmanisi, así que también las habrá en los humanos. Es una lástima que el diente analizado [Welker F et al, 2020] era un trozo muy roto y contaminado. Por supuesto, hay una relación entre H. erectus y H. antecessor, porque comparten un antecesor común. Pero estoy convencido de que hay que ir muy atrás en el tiempo para encontrarlo. Quizá haya que llegar hasta la época de Dmanisi. Lo veremos, seguro.
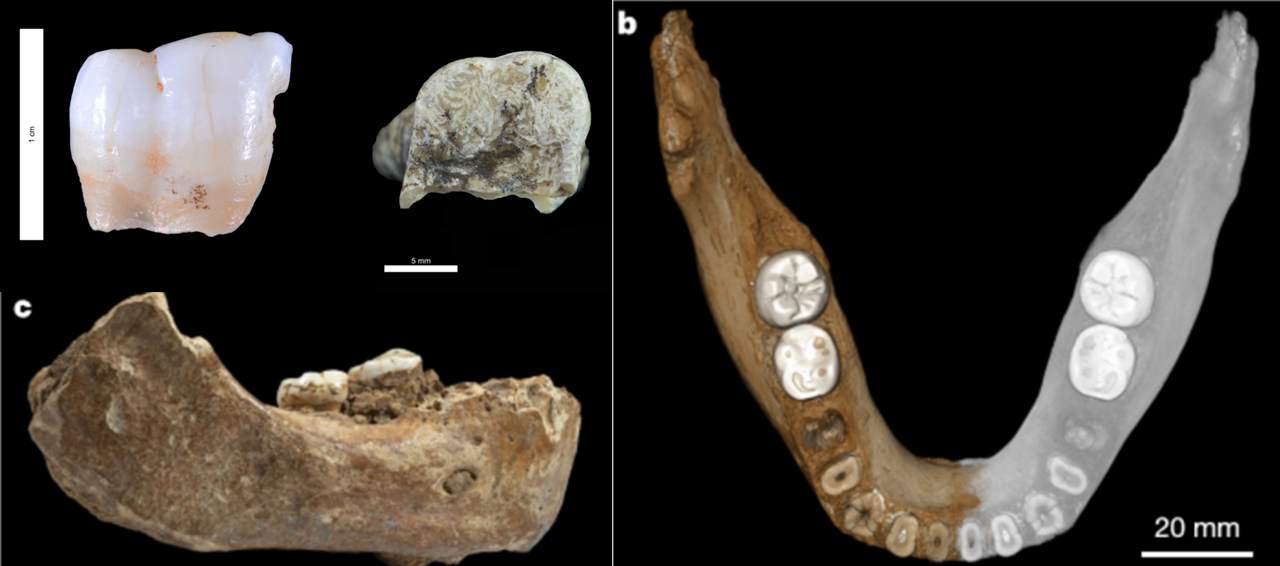
Arriba izda: dientes ATD6-92 de Homo antecessor (Gran Dolina) y D4163 de Homo erectus (Dmanisi). Crédito: Welker F, Ramos-Madrigal J, Gutenbrunner P et al (2020). The dental proteome of Homo antecessor. Nature / Figuras b y c: mandíbula denisovana de Xiahe. Crédito: Chen F. et al (2019). A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau.




