Estuve tentado de dividir esta entrada en varias, ya que en este trimestre hemos conocido un número muy alto de novedades importantes. Finalmente, he reunido y resumido 54 de ellas (!) en un solo artículo que ofrece una visión fascinante de la evolución humana. Espero que lo disfrutéis
- La selección de herramientas de piedra por parte de los chimpancés salvajes para romper nueces en Bossou (Guinea), en experimentos controlados que introducen rocas desconocidas para esta población, podría reflejar las técnicas de los homininos que desarrollaron la industria olduvayense (Braun et al., 2025). Los chimpancés seleccionan yunques y martillos en función de las propiedades mecánicas, características no directamente visibles para el individuo: piedras más duras para los martillos y más blandas para los yunques, lo que indica que comprenden propiedades específicas para funciones distintas. Los adultos identifican las propiedades mecánicas mediante el aprendizaje individual, y los jóvenes a menudo reutilizan las herramientas seleccionadas por los adultos.
- El origen de la talla lítica pudo haber sido un proceso emulativo por parte de los homininos que buscaban reproducir objetos afilados producidos de forma natural («naturalitos») y que ya eran muy demandados, y no tanto la creación intencional o por accidente de lascas afiladas y la posterior aplicación a objetos para cortar (Eren et al., 2025).

Ejemplos de objetos de piedra con filos producidos naturalmente. Crédito: Michelle R. Bebber, Metin I. Eren y Alastair Key
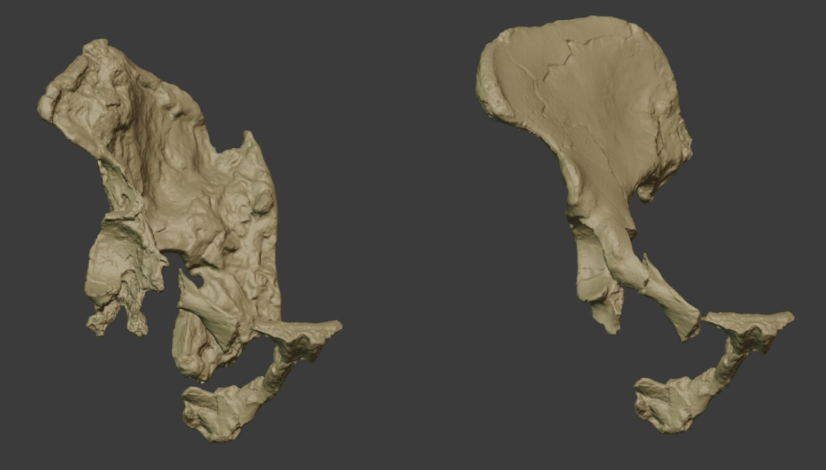
- La orientación acetabular de los Australopithecus era esencialmente similar a la humana hace 3,6 Ma, atendiendo a la comparativa de los acetábulos en una muestra de primates antropoides existentes y homininos fósiles que incluyen Australopithecus afarensis (AL 288–1, KSD-VP-1/1), Australopithecus africanus (Sts 14), Australopithecus sediba (MH2) y Homo neanderthalensis (Kebara 2). El cambio en la orientación acetabular de los homininos fue probablemente un rasgo seleccionado para la locomoción (Lawrence, Hammond, Ward, 2025).
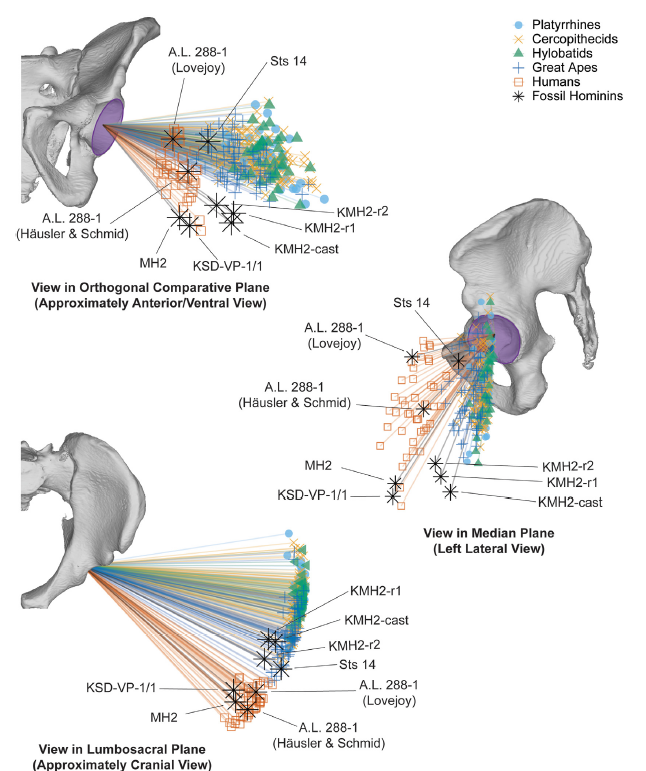
Vectores de orientación acetabular en 3D (vectores normales al plano acetabular) superpuestos en una pelvis humana y mostrados en vistas que representan el plano medio, el plano lumbosacro y el plano comparativo ortogonal. Figura 4 en A.B. Lawrence, A. S. Hammond, C. V. Ward (2025)
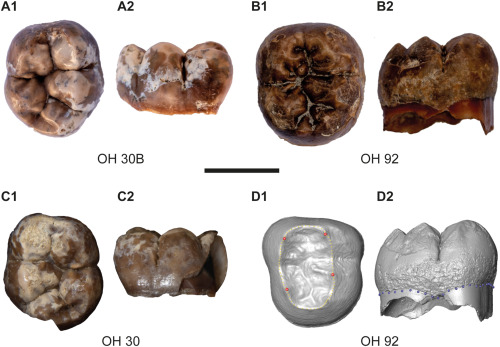
- Se han comparado los isótopos de carbono y nitrógeno unidos al esmalte dental de los dientes fósiles de 43 mamíferos de hace 3,5 Ma de Sterkfontein (Sudáfrica), incluidos 7 individuos diferentes de Au. africanus, con los dientes de mamíferos africanos modernos (Lüdecke et al., 2025). Cuando los animales digieren los alimentos, sus cuerpos tienden a utilizar y eliminar por la orina, las heces y el sudor más el isótopo ligero de nitrógeno (14N) que el isótopo pesado (15N), el cual queda más en sus cuerpos en comparación con los alimentos que comen. Las plantas contienen mucho 14N pero no mucho 15N; los herbívoros tienen más 15N y menos 14N en sus cuerpos que las plantas que comen; y los carnívoros tienen mucho 15N y no mucho 14N. El estudio observa que los australopitecos parecen haber tenido una dieta variada, pero no rica en carne de mamíferos. Este hallazgo podría implicar que las habilidades de los australopitecos para adaptarse a ecosistemas de sabana pudo haber sido anterior al cambio a una dieta rica en carne que potenció la evolución del cerebro. Tampoco se puede descartar una dieta rica en termitas, alimento que contiene relativamente menos nitrógeno del tipo que se encuentra en la carne de los mamíferos.